Bioimpacts. 12(4):349-358.
doi: 10.34172/bi.2021.23571
Original Research
miR-1290 contributes to oncogenesis and angiogenesis via targeting of THBS1, DKK3 and, SCAI
Mohammad Hasan Soheilifar 1  , Majid Pornour 2 , Massoud Saidijam 1, Rezvan Najafi 1, Farid Azizi Jalilian 3, Hoda Keshmiri Neghab 2, Razieh Amini 1, *
, Majid Pornour 2 , Massoud Saidijam 1, Rezvan Najafi 1, Farid Azizi Jalilian 3, Hoda Keshmiri Neghab 2, Razieh Amini 1, *
Author information:
1Research Center for Molecular Medicine, School of Medicine, Hamadan University of Medical Sciences, Hamadan, 6517838678, Iran
2Department of Photo Healing and Regeneration, Medical Laser Research Center, Yara Institute, ACECR, Tehran, 1315795613, Iran
3Department of Virology, Faculty of Medicine, Hamadan University of Medical Sciences, Hamadan, 6517838678, Iran
Abstract
Introduction:
Colorectal cancer (CRC) is the third most common cancer in the world with high mortality, hence, understanding the molecular mechanisms involved in the tumor progression are important for CRC diagnosis and treatment. MicroRNAs (miRNAs) are key gene expression regulators that can function as tumor suppressors or oncogenes in tumor cells, and modulate angiogenesis as a critical process in tumor metastasis. MiR-1290 has been demonstrated as an onco-miRNA in various types of cancer, however, the role of miR-1290 in CRC is not fully understood. This study aimed to investigate the oncogenic and angiogenic potential of miR-1290 in CRC.
Methods:
Lenti-miR-1290 was transduced into HCT116, SW480, and human umbilical vein endothelial cells (HUVECs). By bioinformatics analysis, we identified thrombospondin 1 (THBS1) as a novel predicted target for miR-1290. Quantitative real-time PCR, western blotting, and luciferase reporter assay were used to demonstrate suppression of miR-1290 target genes including THBS1, Dickkopf Wnt signaling pathway inhibitor 3 (DKK3), and suppressor of cancer cell invasion (SCAI) in HCT116 and HUVECs. Cell cycle analysis, proliferation, migration and, tube formation were determined by flow cytometry, MTT, wound healing, and tube formation assays, respectively.
Results:
MiR-1290 significantly decreased the expression of THBS1, DKK3, and SCAI. We demonstrated that miR-1290 enhanced proliferation, migration, and angiogenesis partially through suppression of THBS1, DKK3, and SCAI in CRC.
Conclusion:
These results suggest a novel function of miR-1290 which may contribute to tumorigenesis and angiogenesis in CRC.
Keywords: miR-1290, THBS1, DKK3, SCAI, Oncogenesis, Angiogenesis
Copyright and License Information
© 2022 The Author(s).
This work is published by BioImpacts as an open access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (
http://creativecommons.org/licenses/by-nc/4.0/). Non-commercial uses of the work are permitted, provided the original work is properly cited.
Introduction
miRNAs are small, non-coding RNAs that contribute to gene expression regulation. Their expression can be altered in different types of tumors, including colorectal cancer (CRC). Tumor progression is accompanied by critical mechanisms, including increased proliferation, migration, invasion, impaired regulation of apoptosis, maintenance of cancer stem cells, and angiogenesis. All these mechanisms are involved in increased oncogene expression and/or the downregulation of tumor suppressor genes, which are mediated by miRNA regulatory function.
1
Tumor angiogenesis is one of the main mechanismsregulated by miRNAs and involved in the progression of various cancers, including CRC. Tumor angiogenesis can be caused by miRNA-mediated disruption of the balance between angiogenic factors and their inhibitors.
2
MiR-1290 is an onco-miRNA (oncomiR) that is overexpressed in CRC compared to normal tissues, and it promotes tumor progression.
3
The oncogenic role of miR-1290 has been observed in gastric, lung, ovarian, pancreatic, prostate, esophageal, and breast cancers.
4-11
We have previously shown that miR-1290 is upregulated in CRC tissue compared with a normal margin.
12
The overexpression of miR-1290 via hMSH2 targeting in mismatch repair (MMR)-defective CRC leads to chemotherapy resistance in stage 2 and 3 patients compared to functional MMR tumors.
13
Wu et al have demonstrated that miR-1290 causes cytokine delay and multinucleated cell formation in CRC by targeting kinesin family member 13B (KIF13B).
14
In addition, they have shown that the miR-1290 contributes to reprogramming tumor cells by activating the Wnt pathway and increasing c-Myc and Nanog expression. Furthermore, Nanog, oct4, and sox2 expression could be increased under hypoxic conditions, which is essential for tumor angiogenesis.
15
C- Myc upregulation in colon cancer cells can lead to increased hypoxia-inducible factor 1-alpha (HIF-1α) and vascular endothelial growth factor (VEGF) expression, thereby promoting angiogenesis.
16
Hence, we have investigated whether miR-1290 can act as an angioregulatory miRNA. To the best of our knowledge,the angiogenic potential of miR-1290 has not been studied. We have investigated the regulation of thrombospondin 1 (THBS1), suppressor of cancer cell invasion (SCAI), and Dickkopf Wnt signaling pathway inhibitor 3 (DKK3) by miR-1290. THBS1, as a matrix-bound glycoprotein, is synthesized by various cells, including endothelial cell (EC), and its downregulation is highly associated with increased proliferation and angiogenesis in the murine model of CRC.
17,18
SCAI inhibits the migration and invasion of tumor cells and restricts the development of different types of tumors, including CRC.
19
As far as we know no previous research has investigated the SCAI role in tumor angiogenesis. DKK3 is a tumor suppressor and diagnostic biomarker in CRC,
20
and its inhibition promotes human umbilical vein endothelial cells (HUVECs) proliferation and angiogenesis.
21
The aim of this in vitro study was to evaluate the oncogenic and angiogenic potential of miR-1290 in CRC cells and HUVECs.
Materials and Methods
Bioinformatics analysis
miRNA-target interactions are based on various parameters such as seed match and thermodynamic stability. THBS1, as a potential target of the miR-1290, was screened in miRNA target prediction tools including DIANA-microT web-server v5.0
22
and RNA Hybrid.
23
MiTG is a prediction score in DIANA-microT which is based on conserved and non-conserved miRNA target sites. On the other hand, RNA hybrid is useful for predicting minimal free energy (MFE) in miRNA-target binding.
24
Cell culture
HCT116, SW480, HEK293T, and HUVEC were obtained from the Pasteur Institute (Tehran, Iran). Cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 100 mg/mL streptomycin, and 100 U/mL penicillin (all obtained from Gibco, Paisley, UK) in a humidified incubator (Memmert, Germany) at 37°C with 5% CO2.
MiR-1290 cloning and transduction
MiR-1290 precursors were amplified from human genomic DNA (NG_012283.1) by polymerase chain reaction (PCR). Then, PCR products were cloned into a PEZ-LV105a vector (GeneCopia, Rockville, USA) using XmnI (Sinaclon, Iran), NotI (Sinaclon, Iran) restriction enzymes, and T4 DNA ligase (Sinaclon, Iran). Subsequently, lentivirus was produced from the transformation of the lentivirus-producing vectors, including PEZ-LV105a, packaging plasmid pLV-HELP (InvivoGen, San Diego, CA), and envelope plasmid pMD2G (InvivoGen, San Diego, CA), in HEK293T cells. Viruses were purified and concentrated with Amicon Ultra-4 centrifugal filter units (Millipore, Burlington, USA). Then, the cells were transduced with lenti-miR-1290 based on the manufacturer’s protocol. They were then incubated at 37°C in a humidifier (5% CO2) for 24 hours. Transduced cells were subsequently exposed to 2.5 μg/mL puromycin (Sigma, St. Louis, MO, USA) for two days to obtain stable transductants.
25
Reverse transcription-quantitative polymerase chain reaction (RT-qPCR)
The total RNA of the cells was extracted using a RiboEx LS reagent (Geneall, South Korea), and complementary DNA (cDNA) was synthesized using a first-strand cDNA synthesis kit (Thermo Fisher Scientific, USA). The cDNA samples were amplified by PCR with a panel of specific primers (Table 1 and Table S1). QRT‐PCR was performed to evaluate the expression alterations of the miRNA and its targets using an SYBR Green master mix kit (Genaxxon kit, Germany) with a Mic qPCR cycler (BioMolecular Systems, Australia). At least three biologic replicates of each group were included in the qPCR experiments. β-actin and RNU6B (U6) were used to normalize the quantitative data. QRT‐PCR data were analyzed via the 2 -ΔΔCT method.
Table 1.
The primers sequences used in quantitative RT-PCR and primers for miR-1290 precursor cloning which contain restriction sites for restriction enzymes (Not1 and Xmn1)
|
Gene
|
Forward primer (5′ to 3′)
|
Reverse primer (5′ to 3′)
|
Product length (bp)
|
| Β-actin |
AGACGCAGGATGGCATGGG |
GAGACCTTCAACACCCCAGCC |
161 |
| THBS1 |
CCTGCCATCCGCACTAACTAC |
GTTCTCTTCAGTCACTTTGCGG |
162 |
| DKK3 |
GCACCGAGAAATTCACAAGATAACC |
CTCGTCGATGATGCACTCGT |
121 |
| SCAI |
GCAACTGTTCAATGGGTTAAGAGA |
AAGCCATACCGATTATCCAAGACT |
144 |
| U6 |
CGCTTCGGCAGCACATATACT |
CGCTTCACGAATTTGCGTGTC |
145 |
| miR-1290 |
AGGTGGATTTTTGGATCAGGGA |
GTGCAGGGTCCGAGGT |
64 |
| miR-1290 precursor |
CTAGAAGGAGTTCTGGACTTGCTTGGATTTTGGAGAA |
CTTGCGGCCGCAGGCAACATTTGTCAAGCACTTG |
281 |
Luciferase activity assay
The 3′-UTR of THBS1, DKK3, and SCAI was amplified. Then, the PCR products were inserted into a psiCHECK-2 vector (Promega, USA) using Xho1 (Sinaclon, Iran) and NotI (Sinaclon, Iran). PEZ-LV105a containing miR-1290, an empty psiCHECK-2 (mock), and psiCHECK-2 contained the Renilla luciferase gene upstream of the 3′-UTR target gene. This targeted gene was transfected into the HEK-293T cells seeded in 96-well plates. The relative luciferase activities were evaluated 48 h after transfection using the DualGlo luciferase assay (Promega, USA) according to the manufacturer’s instructions.
25
The primers used for the cloning reactions are shown in Table S2. The predicted binding site on 3’UTR between miR-1290 of THBS1, DKK3, and SCAI respectively are represented in Fig. S1 (See Supplementary file 1).
Western blotting
Proteins were extracted from the untreated, mock, and lenti-miR-1290 groups. Bradford assay was used to normalize the concentration of the extracted samples. 20 μg of each protein sample was subjected to SDS–PAGE in 14% acrylamide gels and transferred to a polyvinylidene difluoride membrane (Millipore, Bedford, MA, USA). A combination of 5% non-fat dry milk and 0.05% Tween-20 was used for blocking and washing in TBS and TBST, respectively. The blots were probed with primary antibodies, including and GAPDH, THBS1, DKK3, and SCAI (Santa Cruz, USA), which were diluted in TBST (1:1000) and incubated for 2 h at room temperature. The membranes were incubated with goat anti-mouse HRP-conjugated secondary IgG for 1 hour at room temperature. Afterward, the immunoreactive bands were detected using an ECL western blotting substrate (ECL, Amersham, Bucking Hampshire, UK).
Colony formation assay
The single-cell suspension was obtained by trypsinization after transduction for 24 hours. 2000 cells were seeded in 60 mm petri dish and cultured for 7 days with DMEM+10%FBS in 5% CO2 incubator. 4% paraformaldehyde and 1% crystal violate dye (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) were used to fix and stain the colonies respectively. Finally, the number of colony-forming units was determined.
Cell proliferation assay
Cellular proliferation was evaluated using an MTT assay. Transduced and non-transduced cells were seeded at 8 × 104 cells/well in 96-well plates. The MTT 3-(4, 5-dimethylth-iazole-2-yl)-2, 5-biphenyl tetrazolium bromide solution (Bioidea, Iran) was added to the medium 24 hours after transduction. Then, cells were incubated for 4 h. Subsequently, a dimethyl sulfoxide solution was added to each well, and the optical density of each was measured with a microplate reader at 570 nm. All tests were performed in triplicate.
Cell cycle analysis by flow cytometry
In this step, 1×106 cells were trypsinized and then washed by phosphate-buffered saline (PBS) (Bioidea, Iran) 72 hours after transduction. Then, the cells were fixed in 70% cold ethanol before being treated in 10 mmol/L RNase (Pishgam biotech co, Iran) at 37°C for 10 minutes. The cells were then stained with 10 μmol/L propidium iodide (Sigma, USA) for 20 minutes in a dark room at 37°C. Afterward, the cell cycle was assessed using FACSCaliburTM flow cytometry (BD Biosciences, San Jose, CA, USA).
Scratch-wound healing assay
Transduced and non-transduced cells were seeded in six-well plates. Upon reaching 90% confluence, a sterile 1000-μL pipette tip was used to scrape the confluent cell layer. Phase-contrast images of scratches were acquired under a microscope (Olympus Corporation, Tokyo, Japan) at 0 and 24 hours. Cells were gently washed with PBS to remove cellular debris and were then allowed to migrate toward the scratched area. Images of the wounds were taken at the same site as the initial image 24 hours after scratching. Wound healing ability was assessed by measuring the alteration of the scraped area using ImageJ software. The percentage of gap healing was calculated using the below equation:
Tube formation assay
The 96-well plates were coated with 10 mg/mL Matrigel (Corning, USA). Transduced and non-transduced HUVECs (2×104/well) were seeded in the Matrigel-coated plates and incubated for 24 hours. Tube-like structures were observed, and three random microscopic fields per well were captured. Images were analyzed using WimTube Image Analysis software (ibidi GmbH, Germany).
Statistical analysis
The results are presented as means ± SEM. One-way ANOVA and t test and were used for statistical comparison. A P value of less than 0.05 (P < 0.05) was considered statistically significant. All tests were repeated three times.
Results
Expression of miR-1290 increased in CRC cell lines and HUVECs after lenti-miR-1290 transduction
miR-1290 expression was increased in HCT116 and SW480 transduced with lenti-miR-1290 compared to the mock vector control group (vector without insert) and untreated group (P < 0.01). The miR-1290 expression level in SW480 was less than that of HCT116 (Figs. 1A and 1B). Due to the higher expression of miR-1290 in HCT116, this cell line was identified as a candidate for further evaluating miR-1290 function. Moreover, miR-1290 expression was upregulated (>3-fold)in lenti-miR-1290 transduced HUVECs compared to mock and untreated cells (P < 0.01) (Fig. 1C).
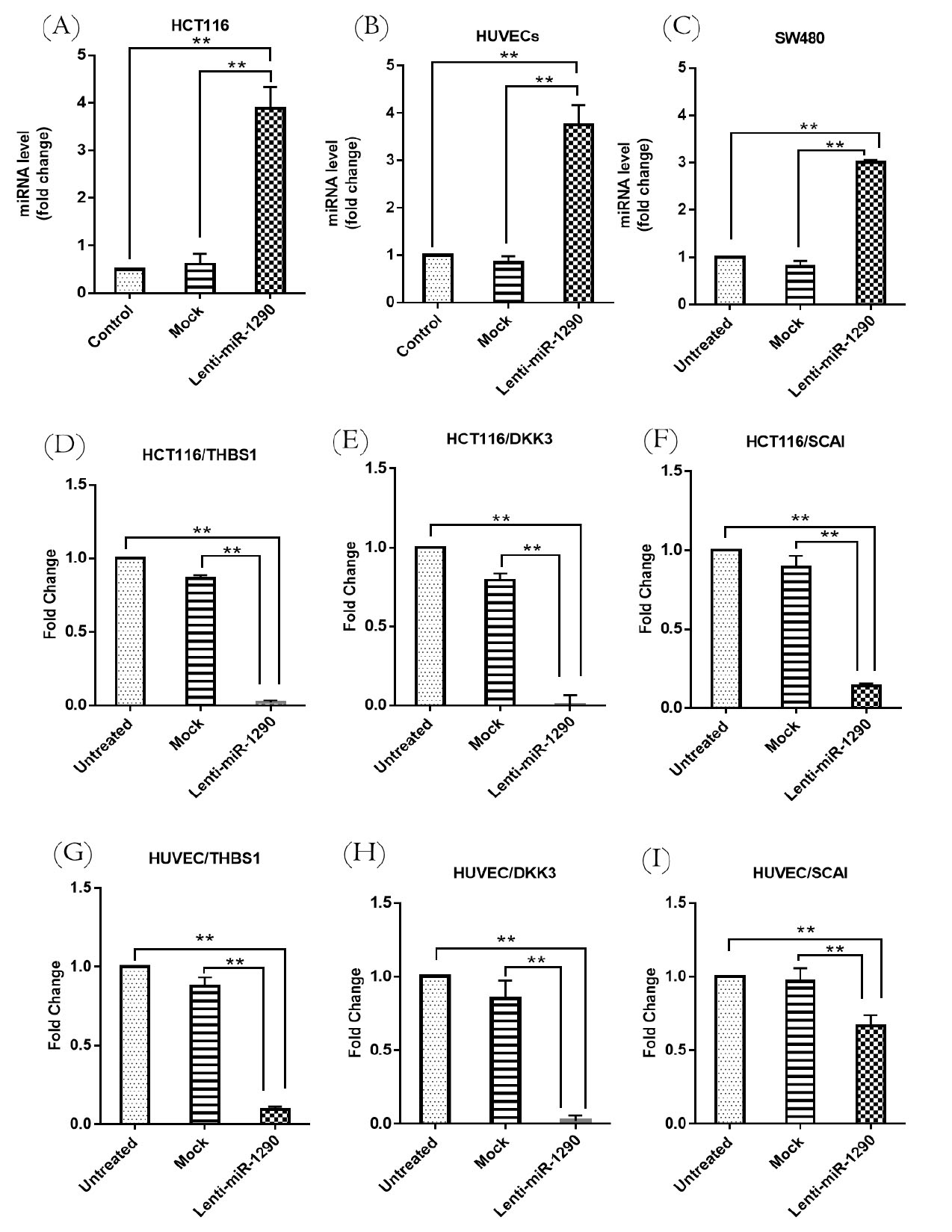
Fig. 1.
Relative expression analysis of miR-1290 and its targets using qRT-PCR after lenti-miR-1290transduction.
Increased expression of miR-1290 in the lenti-miR-1290-treated group compared with untreated and mock groups in HCT116 (A), HUVEC (B), and SW480 (C). THBS1 (D, G), DKK3. (E, H), and SCAI (F, I) were decreased in HCT116 and HUVEC following by miR-1290 transduction. Results are depicted as the mean ± SEM. Each test was repeated three times. ** P < 0.01.
.
Relative expression analysis of miR-1290 and its targets using qRT-PCR after lenti-miR-1290transduction.
Increased expression of miR-1290 in the lenti-miR-1290-treated group compared with untreated and mock groups in HCT116 (A), HUVEC (B), and SW480 (C). THBS1 (D, G), DKK3. (E, H), and SCAI (F, I) were decreased in HCT116 and HUVEC following by miR-1290 transduction. Results are depicted as the mean ± SEM. Each test was repeated three times. ** P < 0.01.
THBS1, DKK3, and SCAI expression suppressed by miR-1290 in HCT116 and HUVEC
The association between miR-1290 expression and its target genes including THBS1 was investigated in HCT116 and HUVECs. THBS1 was predicted as a target of miR-1290 by bioinformatics analysis. miTG score for THBS1 was 0.78 which indicates a high probability of targeting. Moreover, THBS1 was predicted as a putative target in RNA hybrid, and MFE value was -24.1 (kcal/mol). THBS1, as a high score potential target of miR-1290, was selected for further evaluation due to its essential role in angiogenesis and tumor development The purpose of this investigation was to identify the molecular mediators underlying oncogenesis and angiogenesis that might be mediated by miR-1290. The results indicated that THBS1, DKK3, and SCAI were downregulated significantly in HCT116 as a result of miR-1290 upregulation in the lenti-miR-1290 group (P < 0.01) (Fig. 1D-F). THBS1, DKK3, and SCAI were decreased in HUVECs transduced with lenti-miR-1290 in comparison to the mock and untreated groups (P < 0. 01) (Fig. 1G-I). The luciferase activity was decreased in cells co-transfected with miR-1290 and recombinant psiCHECK-2 vectors (containing 3′-UTR of target genes) (P < 0.01) (Fig. 2).
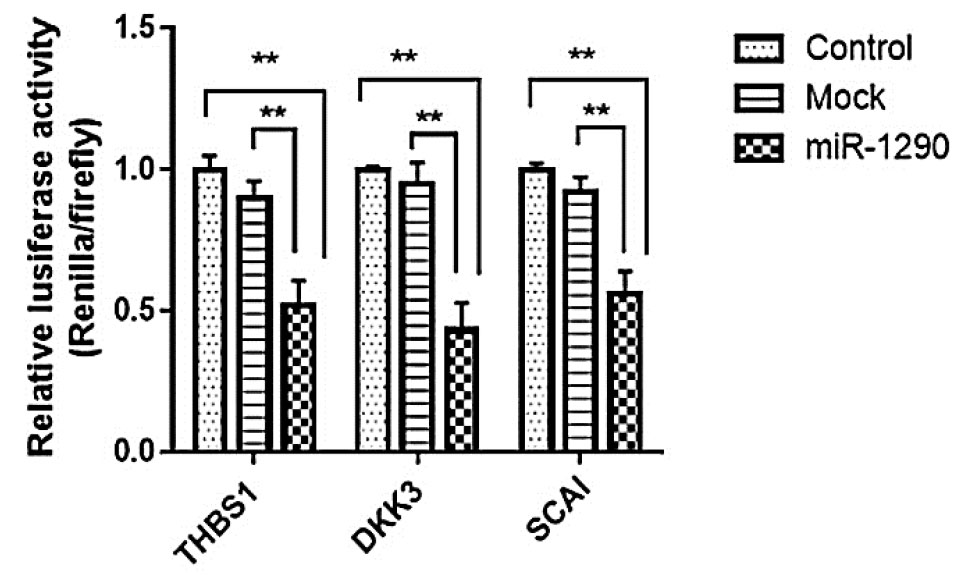
Fig. 2.
THBS1, DKK3 and SACI are direct targets of miR-1290.
Luciferase assay of cells co-transfected with mock control vector (empty psiCHECK-2), miR-1290 or a luciferase reporter constructs containing target genes (THBS1, DKK3 and SCAI) 3′UTR. Control groups are cells transfected with psiCHECK-2 containing 3’UTR without miR-1290 overexpression.
.
THBS1, DKK3 and SACI are direct targets of miR-1290.
Luciferase assay of cells co-transfected with mock control vector (empty psiCHECK-2), miR-1290 or a luciferase reporter constructs containing target genes (THBS1, DKK3 and SCAI) 3′UTR. Control groups are cells transfected with psiCHECK-2 containing 3’UTR without miR-1290 overexpression.
The western blotting analysis confirmed the qRT-PCR results. THBS1 protein levels were reduced in lenti-miR-1290 transduced HCT116 and HUVECs (P < 0.05 and P < 0.01, respectively). Moreover, DKK3 was downregulated in lenti-miR-1290 transduced HCT116 and HUVECs (P < 0.01). The same results were obtained for SCAI suppression in lenti-miR-1290 transduced HCT116 and HUVECs (P < 0.01 and P < 0.05, respectively) (Fig. 3).

Fig. 3.
Western blots results.
The Western blots results of miR-1290 target proteins in HCT116 (A) HUVECs (C) were quantified and the data are shown for miR-1290 overexpressing HCT116 (B) and HUVEC (D). GAPDH was used as an internal control in all the western blot experiments. The results are expressed as the mean ± SEM. *P < 0.05, **P < 0.01,*** P ˂ 0.0001.
.
Western blots results.
The Western blots results of miR-1290 target proteins in HCT116 (A) HUVECs (C) were quantified and the data are shown for miR-1290 overexpressing HCT116 (B) and HUVEC (D). GAPDH was used as an internal control in all the western blot experiments. The results are expressed as the mean ± SEM. *P < 0.05, **P < 0.01,*** P ˂ 0.0001.
MiR-1290 promoted colony forming ability, cell viability, and cell cycle progression in HCT116 and HUVECs
The colony-forming ability of lenti-miR-1290 treated groups showed significant changes compared with mock and untreated groups in both cell lines (Fig. 4A-D) (P < 0.01). Cell viability was significantly increased in the lenti-miR-1290 transduced HCT116 and HUVECs compared with the mock and untreated groups (P < 0.0001 and P < 0.01), respectively (Figs. 4E and 4F). The MTT assay showed that lentivirus had no toxic effect on the cells, as the viability of the mock group was not significantly different from that of the untreated group for either cell line. Flow cytometry analysis demonstrated that miR-1290 upregulation significantly elevated the percentage of cells in the S phase. However, it also reduced the percentage of cells in the G1 phase in comparison to the untreated and mock groups. As such, it appears that miR-1290 can induce cell cycle progression in both cell lines (Fig. 5).
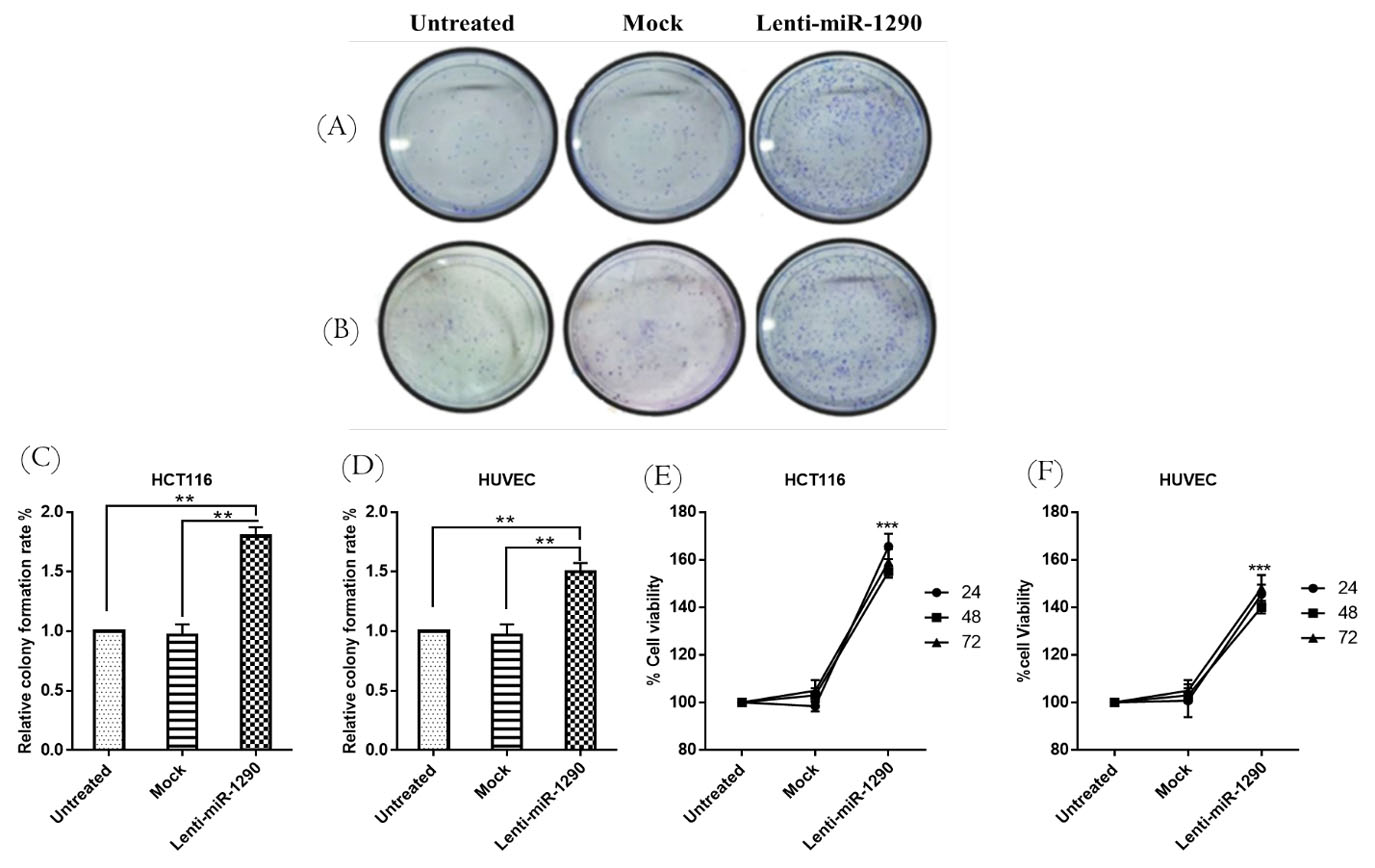
Fig. 4.
Colony formation assay and MTT cell proliferation assay.
The results indicated that lenti-miR-1290 significantly increased colony formation ability in HCT116, and HUVECs compared with untreated and mock (A-D). The rate of cell proliferation was not different between the mock and untreated groups, but cell proliferation was greater in the lenti-miR-1290-treated groups than in the mock and untreated groups in HCT116 (E), and HUVEC (F) at 24 h, 48 h and 72 h. There was no significant difference between different times in the lenti-miR-1290-treated groups. Data are presented as the mean ±SEM of three independent experiments (**P < 0.01).
.
Colony formation assay and MTT cell proliferation assay.
The results indicated that lenti-miR-1290 significantly increased colony formation ability in HCT116, and HUVECs compared with untreated and mock (A-D). The rate of cell proliferation was not different between the mock and untreated groups, but cell proliferation was greater in the lenti-miR-1290-treated groups than in the mock and untreated groups in HCT116 (E), and HUVEC (F) at 24 h, 48 h and 72 h. There was no significant difference between different times in the lenti-miR-1290-treated groups. Data are presented as the mean ±SEM of three independent experiments (**P < 0.01).

Fig. 5.
Cell cycle analysis of HCT116 and HUVECs. Flow cytometry analysis showed that lenti-miR-1290 could induce G1 to Sphaseprogression cell cycle progression in HCT116 (A) and HUVECs (B). Graphical representation of different phases of the cell cycle (G1, S, G2/M). The percentage of cells in G0-G1 was decreased and increased in the S phase compared with mock and control in HCT116 (C) and HUVEC (D). Data are presented as the mean ±SEM of three independent experiments (**P < 0.01).
.
Cell cycle analysis of HCT116 and HUVECs. Flow cytometry analysis showed that lenti-miR-1290 could induce G1 to Sphaseprogression cell cycle progression in HCT116 (A) and HUVECs (B). Graphical representation of different phases of the cell cycle (G1, S, G2/M). The percentage of cells in G0-G1 was decreased and increased in the S phase compared with mock and control in HCT116 (C) and HUVEC (D). Data are presented as the mean ±SEM of three independent experiments (**P < 0.01).
MiR-1290 enhanced migration in HCT116 and HUVECs
The scratch assay results showed that miR-1290 can increase the migration rate of HCT116 and HUVECs. The wound healing ability of both cell lines in the wounded area was higher in the lenti-miR-1290 group than in the mock and untreated groups. There was no difference between the untreated and the mock groups after 24 hours (Fig. 6).
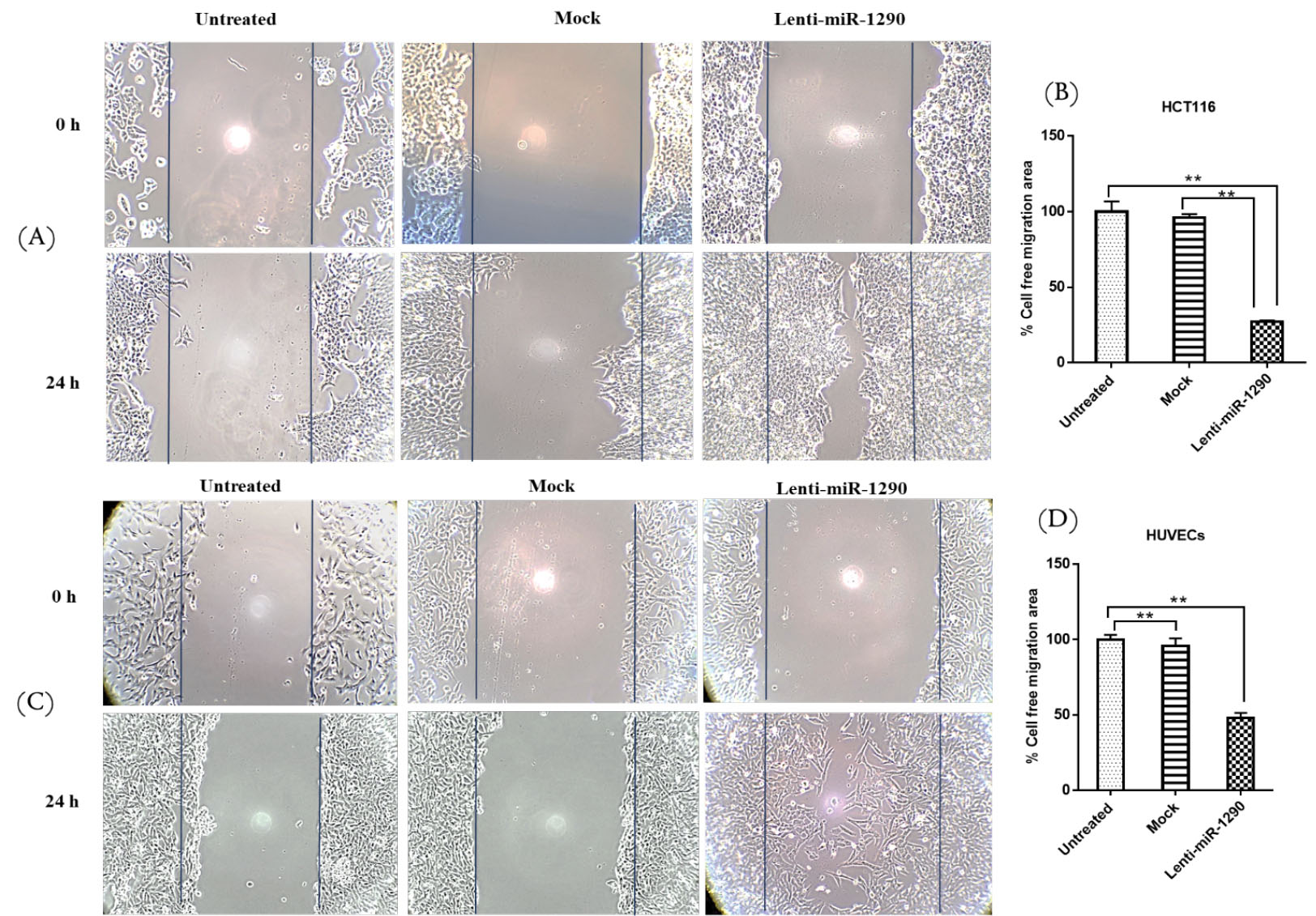
Fig. 6.
Wound scratch assay
. Wound scratch assay for untreated, mock and lenti-miR-1290 treated HCT116 cells (A,B) and HUVECs (C,D). Phase-contrast microscopic images of the wounded area of cells were captured immediately and 24 h after wounding. The images were quantified using the ImageJ software. Data are the mean ± SEM, n = 3, **P < 0.01.
.
Wound scratch assay
. Wound scratch assay for untreated, mock and lenti-miR-1290 treated HCT116 cells (A,B) and HUVECs (C,D). Phase-contrast microscopic images of the wounded area of cells were captured immediately and 24 h after wounding. The images were quantified using the ImageJ software. Data are the mean ± SEM, n = 3, **P < 0.01.
MiR-1290 promoted angiogenesis in HUVECs via THBS1, DKK3, and SCAI suppression
The tube formation assay was performed as an in vitro model of angiogenesis to evaluate the proangiogenic function of miR-1290. The increased expression of miR-1290 in HUVECs transduced with lenti-miR-1290 increased the number of loops and the length of the tubes (Fig. 7).

Fig. 7.
Proangiogenic effect of miR-1290 on the angiogenesis in HUVECs 24 h after transduction. miR-1290 is associated with angiogenesis induction in HUVECs (A, B). Images were analyzed and quantified using WimTube Image Analysis software. Image analysis revealed significant increase of total loops (C), and total tube length (D). *P < 0.05, **P < 0.01.
.
Proangiogenic effect of miR-1290 on the angiogenesis in HUVECs 24 h after transduction. miR-1290 is associated with angiogenesis induction in HUVECs (A, B). Images were analyzed and quantified using WimTube Image Analysis software. Image analysis revealed significant increase of total loops (C), and total tube length (D). *P < 0.05, **P < 0.01.
Discussion
miRNAs are involved in various physiological and pathological processes. They can be incorporated into microvesicles that affect nearby cells by regulating different targets. Several studies have shown the expression regulation of important target genes in angiogenesis and oncogenesis by intracellular and extracellular miRNAs.
2
miRNAs exert tumor-suppressive or oncogenic functions in a context-dependent manner. Angioregulatory miRNAs (angiomiR) are involved in tumorigenesis by modulating the expression of antiangiogenic and proangiogenic factors in tumor cells.
26
They can also trigger paracrine signaling in tumor microenvironments (TME) components, such as ECs, which affect cell proliferation and migration.
27
In addition, endothelial miRNAs can modulate growth factor receptors and signaling molecules in EC. We showed that inhibition of DKK3, as a tumor suppressor by miR-1290, increases proliferation and cell migration in HCT116 cells. The percentage increase of cells in the S phase of the cell cycle in HCT116 cells treated with lenti-miR-1290 is associated with tumor cells’ entry into the proliferative phase. DKK3 induces cell cycle arrest in CRC cell line,
28
so negative regulation of DKK3 by miR-1290 could be partially involved in cell cycle progression. The tumor-suppressive function of DKK3 has also been demonstrated in lung and gallbladder cancers, however, in these cases, it is overexpressed in oral squamous cell carcinoma, which is associated with metastasis and poor prognosis independently of the Wnt pathway.
29,30
Also, β2-microglobulin (β2M) can block DKK3 in ovarian cancer, which activates the VEGFR-2/PI3K/AKT/mTOR pathway.
31
Furthermore, PI3K/AKT dysregulation is involved in CRC angiogenesis.
32
MiR-92a transfection through DKK3 silencing can increase the growth, migration, and tube formation of HUVECs.
33
In the present study, we have shown that angiogenesis was promoted when DKK3 expression was reduced following miR-1290 transduction. Therefore, the antiangiogenic role of DKK3 shown here is consistent with previous studies.
21,34
Furthermore, the scratch assay performed in this study revealed that miR-1290 plays a significant role in cell migration induction. Imaoka et al have shown that miR-1290 suppression restricts CRC cell migration.
35
THBS1 is a potent inducer of prostate tumor cell migration that exerts its effect through CD36, which mediates the antiangiogenic activity of THBS1.
36
Moreover, FGF7/FGFR2 could induce THBS1 expression, which is associated with the invasion and migration of gastric cancer cells.
37
MiR-194 could increase angiogenesis and tumor growth in colon carcinoma xenografts via THBS1 suppression.
38
In the present study, the expression of THBS1 was suppressed by miR-1290 in HCT116. THBS1 suppression increased the migration and proliferation of HCT116 cells, which could be an anti-tumoral factor in CRC cells. We have also shown that the proliferation and migration of HUVECs were increased as a result of miR-1290 overexpression. In addition, the number of loops and the length of tubules increased, suggesting a stimulatory role of miR-1290 in angiogenesis through THBS1 suppression. THBS1 induces cell cycle arrest in ECs,
39
hence, it seems that miR-1290 by targeting THBS1 promotes cell cycle progression in HUVECs.
The results of the present study showed that inhibition of SCAI enhances the proliferation and migration of HCT116 cells. Zheng and colleagues
19
showed that miR-625-3p can reduce CRC cell invasion and migration by inhibiting SCAI. They demonstrated that SCAI suppression could activate MMP9 following E-cadherin downregulation. Another study demonstrated that miR-766-5 can negatively regulate SCAI expression and reduce the proliferation, migration, and invasion of SW480 cells.
40
Reduced SCAI expression leads to Wnt/β-catenin signaling activation in glioma.
41
Because Wnt/β-catenin and MMP9 are involved in angiogenesis,
42
it can be assumed that SCAI might act through this signaling pathway. SCAI downregulation by miR-1290 leads to increased HUVECs proliferation and migration. To the best of our knowledge, no studies have investigated the role of SCAI in angiogenesis inhibition. Furthermore, SCAI through interaction with SWI/SNF complex induces cell cycle arrest.
43
In this study, we showed the progression of ECs cycle followed by negative regulation of SCAI Therefore, it is concluded that miR-1290 induces angiogenesis by stimulating G1 phase progression to S.
Research Highlights
What is the current knowledge?
√ Colorectal cancer (CRC) is the third most common cancer in the world with high mortality.
√ Angioregulatory miRNAs modulate angiogenesis as a critical process in tumor growth and metastasis.
What is new here?
√ Triadic suppression of SCAI, THBS1, and DKK3 by miR-1290 as a novel angiomiR is involved in angiogenesis and oncogenesis.
√ miR-1290 could be considered in determining applicable non-coding RNA-based therapies in future studies.
Conclusion
Altogether, the finding of this in vitro study indicated miR-1290 significantly decreased the expression of THBS1, DKK3, and SCAI. MiR-1290 enhanced proliferation, migration, and angiogenesis partially through triadic suppression of THBS1, DKK3, and SCAI.
Acknowledgments
We would like to acknowledge Mrs. Jahanshiri for her technical support.
Funding sources
This work was funded by the Hamadan University of Medical Sciences.(grant No. 9611247417).
Ethical statement
This work does not include human participants or animal studies.
Competing interests
The authors declare that they have no competing interests.
Authors’ contribution
MHS carried out most of the experiments. MHS and MP designed the experiments and analyzed the data. MHS and R.N participate in drafting and revising the article. FAJ and HKN contributed to data collection. RA and MS critically reviewed the study. All authors reviewed the results and approved the final version of the manuscript.
Supplementary Materials
Supplementary file 1 contains Fig. S1 and Tables S1 & S2.
(pdf)
References
- Thomas J, Ohtsuka M, Pichler M, Ling H. MicroRNAs: Clinical Relevance in Colorectal Cancer. Int J Mol Sci 2015; 16:28063-76. doi: 10.3390/ijms161226080 [Crossref] [ Google Scholar]
- Soheilifar MH, Grusch M, Neghab HK, Amini R, Maadi H, Saidijam M. Angioregulatory microRNAs in Colorectal Cancer. Cancers (Basel) 2019; 12:1-20. doi: 10.3390/cancers12010071 [Crossref] [ Google Scholar]
- Ma Q, Wang Y, Zhang H, Wang F. miR-1290 Contributes to Colorectal Cancer Cell Proliferation by Targeting INPP4B. Oncol Res 2018; 26:1167-74. doi: 10.3727/096504017x15051741798389 [Crossref] [ Google Scholar]
- Li A, Yu J, Wolfgang C, Hruban R, Goggins M, Kim H. MicroRNA Array Analysis Finds Elevated Serum miR-1290 Accurately Distinguishes Patients with Low-Stage Pancreatic Cancer from Healthy and Disease Controls. Clin Cancer Res 2013; 19:3600-10. doi: 10.1158/1078-0432.CCR-12-3092 [Crossref] [ Google Scholar]
- Kim G, An H-J, Lee M-J, Song J-Y, Jeong J-Y, Lee J-H. Hsa-miR-1246 and hsa-miR-1290 are associated with stemness and invasiveness of non-small cell lung cancer. Lung Cancer 2016; 91:15-22. doi: 10.1016/j.lungcan.2015.11.013 [Crossref] [ Google Scholar]
- Kobayashi M, Sawada K, Nakamura K, Yoshimura A, Miyamoto M, Shimizu A. Exosomal miR-1290 is a potential biomarker of high-grade serous ovarian carcinoma and can discriminate patients from those with malignancies of other histological types. J. Ovarian Res 2018; 11:81. doi: 10.1186/s13048-018-0458-0 [Crossref] [ Google Scholar]
-
Mao Y, Liu J, Zhang D, Li B. MiR-1290 promotes cancer progression by targeting nuclear factor I/X(NFIX) in esophageal squamous cell carcinoma (ESCC). Biomed Pharmacother 2015; 76: 82-93. 10.1016/j.biopha.2015.10.005
- Huang X, Yuan T, Liang M, Du M, Xia S, Dittmar R. Exosomal miR-1290 and miR-375 as Prognostic Markers in Castration-resistant Prostate Cancer. Eur Urol 2015; 67:33-41. doi: 10.1016/j.eururo.2014.07.035 [Crossref] [ Google Scholar]
- Li M, He X-Y, Zhang Z-M, Li S, Ren L-H, Cao R-S. MicroRNA-1290 promotes esophageal squamous cell carcinoma cell proliferation and metastasis. World J Gastroenterol 2015; 21:3245-55. doi: 10.3748/wjg.v21.i11.3245 [Crossref] [ Google Scholar]
-
Aziz F, Yulin L, Yan Q. miR-1290 stimulates proliferation of gastric cancer by targeting SP1 with overexpression of fucosyltransferase IV and α1, 3-fucosylated glycans. bioRxiv 2020; 2020.01.05.895300. 10.1101/2020.01.05.895300
- Endo Y, Toyama T, Takahashi S, Yoshimoto N, Iwasa M, Asano T. miR-1290 and its potential targets are associated with characteristics of estrogen receptor α-positive breast cancer. Endocr Relat Cancer 2013; 20:91-102. [ Google Scholar]
- Nobari S, Soheilifar MH, Neghab HK, Jalilian FA, Amini R. Expression of miR-1290 and Its Target Genes THBS1 and DKK3 in Colorectal Cancer Patients. Basic & Clinical Cancer Research 2020; 12:10-17. doi: 10.18502/bccr.v12i1.5729 [Crossref] [ Google Scholar]
- Ye L, Jiang T, Shao H, Zhong L, Wang Z, Liu Y. miR-1290 Is a Biomarker in DNA-Mismatch-Repair-Deficient Colon Cancer and Promotes Resistance to 5-Fluorouracil by Directly Targeting hMSH2. Mol Ther Nucleic Acids 2017; 7:453-64. doi: 10.1016/j.omtn.2017.05.006 [Crossref] [ Google Scholar]
- Wu J, Ji X, Zhu L, Jiang Q, Wen Z, Xu S. Up-regulation of microRNA-1290 impairs cytokinesis and affects the reprogramming of colon cancer cells. Cancer Lett 2013; 329:155-63. doi: 10.1016/j.canlet.2012.10.038 [Crossref] [ Google Scholar]
- Gawlik-Rzemieniewska N, Bednarek I. The role of NANOG transcriptional factor in the development of malignant phenotype of cancer cells. Cancer Biol Ther 2016; 17:1-10. doi: 10.1080/15384047.2015.1121348 [Crossref] [ Google Scholar]
- Chen C, Cai S, Wang G, Cao X, Yang X, Luo X. c-Myc enhances colon cancer cell-mediated angiogenesis through the regulation of HIF-1α. Biochem Biophys Res Commun 2013; 430:505-11. doi: 10.1016/j.bbrc.2012.12.006 [Crossref] [ Google Scholar]
-
Huang T, Sun L, Yuan X, Qiu H. Thrombospondin-1 is a multifaceted player in tumor progression. Oncotarget 2015; 8. 10.18632/oncotarget.19165
- Lopez-Dee ZP, Chittur SV, Patel H, Chinikaylo A, Lippert B, Patel B. Thrombospondin-1 in a Murine Model of Colorectal Carcinogenesis. PloS One 2015; 10:e0139918-e. doi: 10.1371/journal.pone.0139918 [Crossref] [ Google Scholar]
- Zheng H, Ma R, Wang Q, Zhang P, Li D, Wang Q. MiR-625-3p promotes cell migration and invasion via inhibition of SCAI in colorectal carcinoma cells. Oncotarget 2015; 6:27805-15. doi: 10.18632/oncotarget.4738 [Crossref] [ Google Scholar]
- Safari E, Mosayebi G, Khorram S. Dkk-3 as a potential biomarker for diagnosis and prognosis of colorectal cancer. Med J Islam Repub Iran 2018; 32:86. doi: 10.14196/mjiri.32.86 [Crossref] [ Google Scholar]
- Busceti CL, Marchitti S, Bianchi F, Di Pietro P, Riozzi B, Stanzione R. Dickkopf-3 Upregulates VEGF in Cultured Human Endothelial Cells by Activating Activin Receptor-Like Kinase 1 (ALK1) Pathway. Front Pharmacol 2017; 8:111. doi: 10.3389/fphar.2017.00111 [Crossref] [ Google Scholar]
- Paraskevopoulou MD, Georgakilas G, Kostoulas N, Vlachos IS, Vergoulis T, Reczko M. DIANA-microT web server v5.0: service integration into miRNA functional analysis workflows. Nucleic Acids Res 2013; 41:W169-W73. doi: 10.1093/nar/gkt393 [Crossref] [ Google Scholar]
- Rehmsmeier M, Steffen P, Höchsmann M, Giegerich R. Fast and effective prediction of microRNA/target duplexes. RNA 2004; 10:1507-17. doi: 10.1261/rna.5248604 [Crossref] [ Google Scholar]
- Soheilifar MH, Moshtaghian A, Amini R, Asefi M, Basiri P, Saidijam M. BMI1 as a Potential Target of miR-330-3p in Colorectal Cancer. Middle East J Rehabil Health Stud 2018; 5:1-6. doi: 10.5812/mejrh.66075 [Crossref] [ Google Scholar]
- Soheilifar MH, Vaseghi H, Seif F, Ariana M, Ghorbanifar S, Habibi N. Concomitant overexpression of mir-182-5p and mir-182-3p raises the possibility of IL-17-producing Treg formation in breast cancer by targeting CD3d, ITK, FOXO1, and NFATs: A meta-analysis and experimental study. Cancer Sci 2021; 112:589-603. doi: 10.1111/cas.14764 [Crossref] [ Google Scholar]
- Chen S, Xue Y, Wu X, Le C, Bhutkar A, Bell EL. Global microRNA depletion suppresses tumor angiogenesis. Genes Dev 2014; 28:1054-67. doi: 10.1101/gad.239681.114 [Crossref] [ Google Scholar]
- Kim DY, Lee S-S, Bae Y-K. Colorectal cancer cells differentially impact migration and microRNA expression in endothelial cells. Oncol Lett 2019; 18:6361-70. doi: 10.3892/ol.2019.11055 [Crossref] [ Google Scholar]
- Yang Z-R, Dong W-G, Lei X-F, Liu M, Liu Q-S. Overexpression of Dickkopf-3 induces apoptosis through mitochondrial pathway in human colon cancer. World J Gastroenterol 2012; 18:1590-601. doi: 10.3748/wjg.v18.i14.1590 [Crossref] [ Google Scholar]
- Gondkar K, Patel K, Patil Okaly GV, Nair B, Pandey A, Gowda H. Dickkopf Homolog 3 (DKK3) Acts as a Potential Tumor Suppressor in Gallbladder Cancer. Front Oncol 2019; 9:1121. doi: 10.3389/fonc.2019.01121 [Crossref] [ Google Scholar]
-
Katase N, Lefeuvre M, Tsujigiwa H, Fujii M, Ito S, Tamamura R, et al. Knockdown of Dkk-3 decreases cancer cell migration and invasion independently of the Wnt pathways in oral squamous cell carcinoma-derived cells. Oncol Rep 2013; 29. 10.3892/or.2013.2251
- Kim B-R, Lee E-J, Seo S, Lee S-H, Rho S. Dickkopf-3 (DKK-3) obstructs VEGFR-2/Akt/mTOR signaling cascade by interacting of β2-microglobulin (β2M) in ovarian tumorigenesis. Cell Signal 2015; 27:2150-9. doi: 10.1016/j.cellsig.2015.08.008 [Crossref] [ Google Scholar]
- Sun D, Zhang F, Qian J, Shen W, Fan H, Tan J. 4′-hydroxywogonin inhibits colorectal cancer angiogenesis by disrupting PI3K/AKT signaling. Chem Biol Interact 2018; 296:26-33. doi: 10.1016/j.cbi.2018.09.003 [Crossref] [ Google Scholar]
- Yamada N, Nakagawa Y, Tsujimura N, Kumazaki M, Noguchi S, Mori T. Role of Intracellular and Extracellular MicroRNA-92a in Colorectal Cancer. Transl Oncol 2013; 6:482-92. doi: 10.1593/tlo.13280 [Crossref] [ Google Scholar]
- Ghaffar S, Guo Y, McQueen P, Christ A, Tu K, Xie J. Abstract 3963: Wnt-Signaling regulates angiogenesis by regulating Neuropilin-1 expression in osteosarcoma. Cancer Res 2010; 70:3963. doi: 10.1158/1538-7445.am10-3963 [Crossref] [ Google Scholar]
- Imaoka H, Toiyama Y, Fujikawa H, Hiro J, Saigusa S, Tanaka K. Circulating microRNA-1290 as a novel diagnostic and prognostic biomarker in human colorectal cancer. Ann Oncol 2016; 27:1879-86. doi: 10.1093/annonc/mdw279 [Crossref] [ Google Scholar]
- Firlej V, Mathieu J, Gilbert C, Lemonnier L, Nakhle J, Gallou-Kabani C. Thrombospondin-1 triggers cell migration and development of advanced prostate tumors. Cancer Res 2011; 71:7649-58. doi: 10.1158/0008-5472.CAN-11-0833 [Crossref] [ Google Scholar]
- Huang T, Wang L, Liu D, Li P, Xiong H, Zhuang L. FGF7/FGFR2 signal promotes invasion and migration in human gastric cancer through upregulation of thrombospondin-1. Int J Oncol 2017; 50:1501-12. doi: 10.3892/ijo.2017.3927 [Crossref] [ Google Scholar]
- Sundaram P, Hultine S, Smith LM, Dews M, Fox JL, Biyashev D. p53-responsive miR-194 inhibits thrombospondin-1 and promotes angiogenesis in colon cancers. Cancer Res 2011; 71:7490-501. doi: 10.1158/0008-5472.can-11-1124 [Crossref] [ Google Scholar]
- Oganesian A, Armstrong LC, Migliorini MM, Strickland DK, Bornstein P. Thrombospondins use the VLDL receptor and a nonapoptotic pathway to inhibit cell division in microvascular endothelial cells. Mol Biol Cell 2008; 19:563-71. doi: 10.1091/mbc.e07-07-0649 [Crossref] [ Google Scholar]
- Jia B, Xia L, Cao F. The role of miR-766-5p in cell migration and invasion in colorectal cancer. Exp Ther Med 2018; 15:2569-74. doi: 10.3892/etm.2018.5716 [Crossref] [ Google Scholar]
- Chen X, Hu W, Xie B, Gao H, Xu C, Chen J. Downregulation of SCAI enhances glioma cell invasion and stem cell like phenotype by activating Wnt/β-catenin signaling. Biochem Biophys Res Commun 2014; 448:206-11. doi: 10.1016/j.bbrc.2014.04.098 [Crossref] [ Google Scholar]
- Wu C, Chen J, Chen C, Wang W, Wen L, Gao K. Wnt/β-catenin coupled with HIF-1α/VEGF signaling pathways involved in galangin neurovascular unit protection from focal cerebral ischemia. Sci Rep 2015; 5:16151. doi: 10.1038/srep16151 [Crossref] [ Google Scholar]
- Kreßner C, Nollau P, Grosse R, Brandt DT. Functional interaction of SCAI with the SWI/SNF complex for transcription and tumor cell invasion. PloS One 2013; 8:e69947-e. doi: 10.1371/journal.pone.0069947 [Crossref] [ Google Scholar]