Bioimpacts. 16:30303.
doi: 10.34172/bi.30303
Original Article
Synergistic effects of phytochemical cocktail and aerobic exercise on mRNA and lncRNA networks modulating bone turnover in hypothyroid mice
Malihe Bakhti Conceptualization, Investigation, Methodology, Visualization, Writing – original draft, 1 
Farzaneh Taghian Conceptualization, Investigation, Methodology, Project administration, Supervision, Visualization, Writing – original draft, 1, *
Khosro Jalali Dehkordi Investigation, Validation, Writing – original draft, Writing – review & editing, 1
Rezvan Mirsafaei Rizi Investigation, Validation, Writing – original draft, Writing – review & editing, 2
Author information:
1Department of Sports Physiology, Isf.C., Islamic Azad University, Isfahan, Iran
2Department of Sports injuries, Isf.C., Islamic Azad University, Isfahan, Iran
Abstract
Introduction:
Metabolic bone disease (MBD) is a multifactorial condition that leads to microarchitectural and bone mass disturbance. The association and mechanisms between MBD and hypothyroidism are not comprehensively elucidated. Moreover, long non-coding RNAs (lncRNAs), as critical regulators in biological processes, are one category of RNAs that could regulate gene expression through various mechanisms.
Methods:
Artificial intelligence indicated several mRNAs-lncRNAs involved in the bone tissue in hypothyroidism conditions. After in-silico and chemoinformatic analysis, 30 female C57BL/6 mice at six weeks old and an approximate weight of 24±2 g were divided into five subgroups (n=6 mice/groups), including 1) hypothyroidism in mice (HYPO), 2) hypothyroidism in mice treated with Dorema aucheri (DA) extract (0.4 mg/kg), 3) hypothyroidism in mice treated with aerobic exercise (AE), 4) hypothyroidism in mice treated DA extract bioactive compound along with AE, 5) Normal group (without any induction and treatments). Real-Time PCR detected the relative expression of mRNA and lncRNAs.
Results:
We indicated that the osteoblast and osteoclast markers are dysregulated in hypothyroidism. In addition, DA and AE modified the bone remodeling, bone resorption markers, and inhibitors of Wnt/beta-catenin. We revealed that Adipoq, Dnmt1, Sirt-1, Ppar-ɑ, Ncor1, and Ctnnb1 networks are dysregulated in hypothyroidism conditions. DA and AE modulated this network in bone tissue. The relative expression of the four lncRNAs (TUG1, HOTAIR, H19, and MEG3) was altered in hypothyroidism. Notably, DA and AE significantly regulated the expression level of these lncRNAs.
Conclusion:
AE and DA complementary and alternative medicine ameliorated the metabolic bone disease induced by hypothyroidism in mice.
Graphical Abstract
Keywords: Herbal medicine, Exercise, Rehabilitation, Bioinformatic, Sirt-1
Copyright and License Information
© 2026 The Author(s).
This work is published by BioImpacts as an open access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (
http://creativecommons.org/licenses/by-nc/4.0/). Non-commercial uses of the work are permitted, provided the original work is properly cited.
Funding Statement
There was no funding to support this study.
Introduction
Metabolic bone disease is an essential worldwide problem in which patients suffer from walking and impaired physical ability, resulting in immobility and enhancing the risk of fracture. The dysregulation of metabolic skeletal is a multifactorial status, recognized by disrupting the microarchitectural bone structure and bone mass. Vegetative states, elderly, para- or hemiplegia after spinal cord injury, and thyroid hormone disorders are major risk factors in bone pathogenesis that impair trabecular bone microarchitecture and impaired cortical mineral density.1 Based on epidemiology evidence, there is an association between dysfunction of bone turnover and hypothyroidism, which may influence osteoclast bone-resorbing and osteoblastic bone-forming.2 Furthermore, thyroid hormones can be indispensable in energy expenditure, metabolism, bone homeostasis development, and skeletal system growth.3 On the other hand, thyroid hormones such as Triiodothyronine (T3), Thyroxine (T4), and Thyroid-stimulating hormones (TSH) might regulate the osteoclast and osteoblast generation via thyroid hormone receptor α (TRα).4 Overwhelming evidence has proved that hypothyroidism is caused by enhanced duration of bone remodeling and increased osteoclast activity, which can markedly increase bone fracture risk.5
Through increased expression of osteocalcin (OSX), collagen, and alkaline phosphatase (ALP), THs encourage osteoblast development and bone matrix formation.2 Adults with THs disturbances consisting of hypothyroidism and hyperthyroidism were predisposed to bone fragility and fractures via different mechanisms.5 Hypothyroidism could delay the remodeling cycle, resulting in slower bone turnover and more mineralization.6 Moreover, population studies have indicated that hypothyroidism might have a greater fracture risk, indicating poor bone quality.7 On the other hand, animal studies have indicated that hypothyroidism could stimulate bone density in Trabecular bone, while Cortical bone structure in hypothyroidism conditions remains unchanged.5
The crucial cellular processes, including proliferation, differentiation, cell growth, genetic stability, tissue homeostasis, and apoptosis, are regulated by a highly conserved Wnt/beta-catenin signaling pathway.6,8 The Wnt signaling pathway significantly controls osteoblastogenesis, bone formation, and homeostasis. Hence, the Wnt/beta-catenin signaling pathway is bone biology's most crucial canonical signaling.9 Furthermore, previous analysis predicts that THs interact with the Wnt signalling system locally to regulate terminal differentiation of the growth plate chondrocyte through growth hormones/receptors mechanisms in the molecular docking in PyRx results.10 O'Shea and colleagues found that T3 could suppress Wnt signaling based on the in vitro study Therefore, based on this study, there is a correlation between Wnt signaling and THs.11
Non-coding RNAs such as long non-coding RNAs (lncRNAs) are critical regulators in several biological processes that dysregulate pathogenic status.12 lncRNAs are one category of non-coding RNAs longer than 200 nucleotides in mature structures, and interaction with RNAs, DNAs, epigenetics factors, and proteins could be managed gene expression by various mechanism pathways in transcriptional, post-transcriptional, translational, post-translational, and epigenetically levels.13 Thus, different roles are defined for lncRNAs in gene expression regulation, such as chromatin remodeling, sponge, scaffolding, RNA processing, mRNA translation, mRNA degradation, mRNA splicing modulation, and transcription activation/suppression.14 The immense evidence has shown that lncRNAs are operative in bone remodeling and regulating skeletal tissue homeostasis.
Phytochemical products are beneficial substances with various applications in everyday life. These natural products might be complementary and alternative medicine in preventing and alleviating illness hallmarks synchronized with ordained medicine.15 Based on a recent study, the essential oil of aerial parts of Dorema aucheri (DA) is abundant in flavonoids and terpenoids. Hence, these herbs have antioxidant, anti-tumor, anti-diabetic, anti-hypercholesterolemia, anti-hyperlipidemia, and anti-inflammatory properties.16 Conversely, growing evidence reported that a phytochemical cocktail affected thyroid hormones, sex hormones, and antioxidant enzymes.17 Since safety and effectiveness are essential in herbal medicine administration,.18,19 the effectiveness of the phytochemicals cocktail in this study is estimated based on a computational photochemistry survey over the potential cutpoints in pathogenesis.20 Complementary health approaches defined physical activity and exercise training as effective therapeutic strategies.21 Regular exercise training in the long term could significantly affect body performance, metabolism, and hormone activity.22 Previous studies have shown that alterations in thyroid hormone levels during long-term exercise training led to no change or amplified THs hormone levels.23 Accordingly, this concept is emphasized for thyroid hormone disruption therapy and is defined as a complementary strategy without side effects and functional medicine. Therefore, this study addressed an association between hypothyroidism and metabolic bone disease based on vital Long non-coding RNAs-mRNA interaction and detected the pathomechanism, hub genes, and long-coding RNA involved in this condition. Moreover, aerobic exercise (AE) and consumption of a safe dosage of phytochemicals cocktail could improve hypothyroidism and metabolic bone disease.
Materials and Methods
Study design
Thirty female C57BL/6 mice at six weeks old and an approximate weight of 24 ± 2 g were purchased from Royan institute and were maintained in the animal house of Isfahan (Khorasgan) Branch of Islamic Azad University, under standard condition (23 ± 4°C, 60% ( ± 4%) of humidity and 12 h dark/12 h light cycle (lights from 08:00 am to 8:00 pm). After one week of adaptation, mice were divided into two groups as the control group (without inducing hypothyroidism, without prescribed Phytochemicals cocktail, and untrained). Mice were injected with 8 mg/kg propylthiouracil (PTU) daily for one month to induce hypothyroidism.24 After that, mice were divided into four subgroups, including 1. hypothyroidism in mice (HYPO), 2. hypothyroidism in mice treated with Phytochemicals cocktail (0.4 mg/kg) (HYPO-DA), 3. hypothyroidism in mice treated with aerobic exercise (HYPO-AE), 4. hypothyroidism in mice treated Phytochemicals cocktail bioactive compound along with aerobic exercise (HYPO-DA + AE), (n = 6 mice/groups). They were housed in clean polypropylene cages under standard conditions of humidity (50 ± 5%), temperature (25 ± 2 °C), and light (12 h light/12 h dark cycle) and fed with a standard diet and drinking water ad libitum. Furthermore, the body weight, fat mass, lean mass, and calorie intake were monitored weekly. After the last endurance training and administrated phytochemicals cocktail, the mice were fasted for 6 hours and euthanized under the combined injection of ketamine (80 mg/kg) and xylazine (10 mg/kg). Bone tissues were collected and snap-frozen in Liquid nitrogen. In addition, blood samples were collected, and the serums were separated. Bone tissue and serum samples were transferred to the medical genetics laboratory for storage at -80°C. The workflow of the study is indicated in Fig. 1.
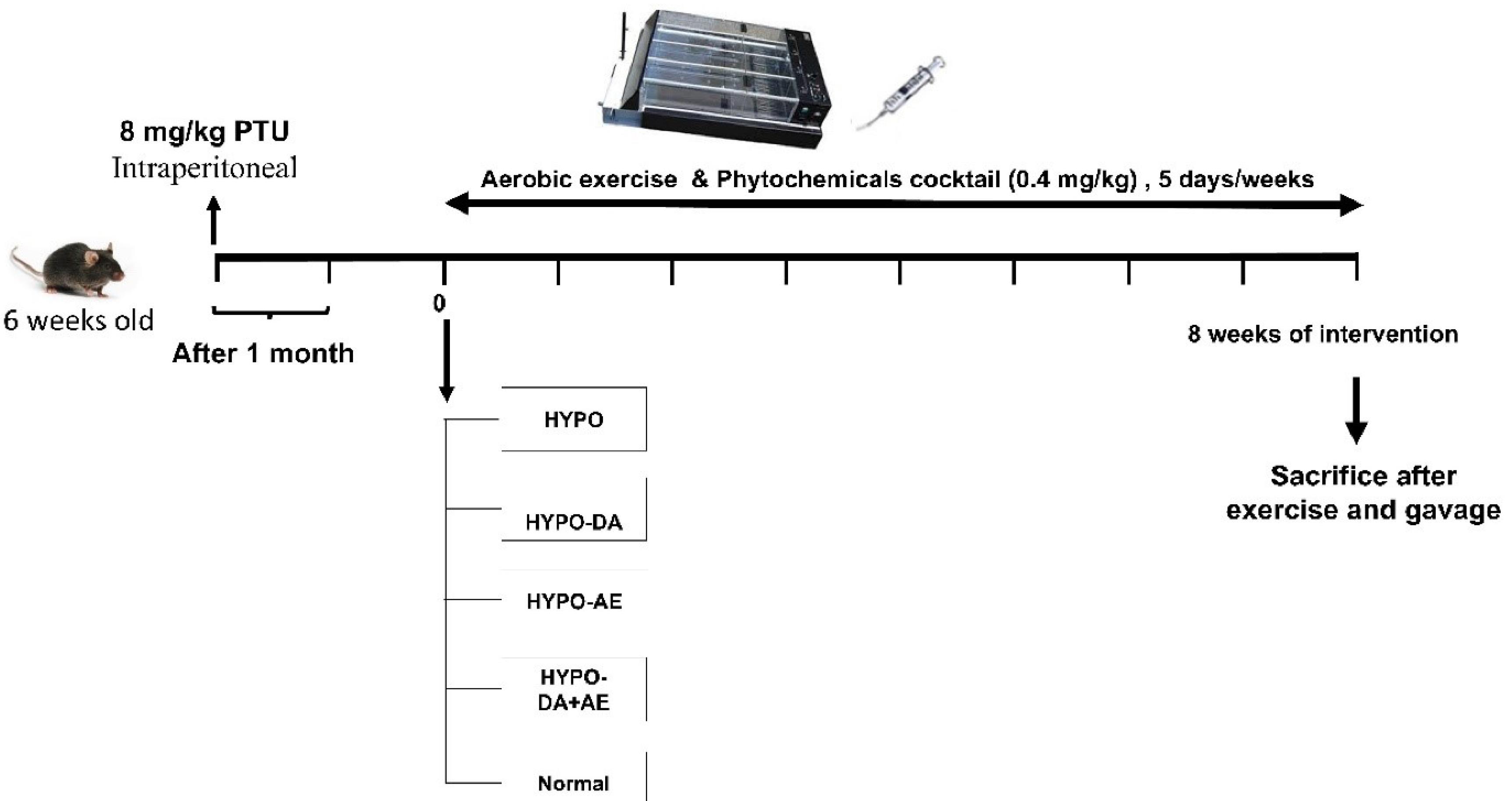
Fig. 1.
Workflow of the study.
.
Workflow of the study.
Gene expression analysis in hypothyroidism and bone metabolism: identifying hub genes and pathways by bioinformatic approaches
We browsed the closest gene expression omnibus microarray dataset to constitute a list of significant genes with differential expression in hypothyroidism status and effect on bone formation and microarchitecture. In this survey, we selected GSE56815 to analyze variable gene expression in bone metabolism, considering low and high bone density in premenopausal women.25 In this integrative analysis, we applied the Specialized packages in Bioconductor, such as hgu133a.db, limma, gcrma, Biobase, simpleaffy, and gplots for identifying differentially expressed Genes. Hence, we conducted microarray dataset analysis in R statistical programming language software and showed the outputs in a heatmap diagram with a significant expression score of P value < 0.001.26 Significant genes with differential expression marked by P value adj < 0.05. Here, we determined the up-regulation and down-regulation of significant genes via logFC threshold + 0.2. We designed a comprehensive network of upregulated and downregulated genes in STRING version 11.5 server based on evidence and medium confidence (0.4) (30) to highlight hub genes involved in bone metabolism disorders. Furthermore, we applied degree = 10, betweenness centrality = 0.005, and closeness centrality = 0.2 to visualize network parameters in Cytoscape software version 3.6.0.27 and marked hub genes in this network. We also classified the hub genes based on function in pathways and designed a network that considered degree, betweenness, and modularity in Gephi 0.9.2 software.28
On the other hand, we collected a gene list associated with hypothyroidism (CUI: C0020676) from the DisGeNET database with a score of gda > 0.01.29 In the following, we schemed the protein-protein interactions (PPIs) network of proteins involved in hypothyroidism conditions. Visualizing network parameters was performed by applying degree = 4, betweenness centrality = 0.004, and closeness centrality = 0.25 in Cytoscape software and highlighting the hub genes involved in hypothyroidism based on evidence algorithm.27 Also, we classified hub genes based on function in pathways in Gephi software.28
Then, we found collaborated hub genes between hypothyroidism and metabolic bone diseases by the VENN diagram bioinformatics tool.30 In the next stage, we constructed a PPIs network for common hub genes between hypothyroidism and metabolic bone diseases to specify critical genes and cutpoint proteins. We marked genes with the highest betweenness centrality and the most degree in this network as the target protein for therapeutic approaches.
Moreover, enrichment and data mining of common genes highlighted the significant molecular signaling pathways involved in metabolic bone pathogenesis, based on Panther and KOBAS-i enrichment databases.31,32 we also browsed the association between hub genes and diseases via the GWAS catalog database.33
Identification and prediction of lncRNAs in metabolic bone diseases
Since lncRNAs are potential genetic molecules in regulating various biological processes such as transcription, gene expression, and mRNA stability, this study aimed to determine the role of lncRNAs in the pathogenesis of metabolic bone disorders hypothyroidism status by measuring expression levels of lncRNAs in mice with hypothyroidism-like. Accordingly, we predicted and identified lncRNAs associated with common genes, especially CTNNB1, as the cut point in the pathogenesis network based on LncTarD.34 LncRNA Disease version 2.0,35 MNDR version 3.1,36 LncBook,37 LNCipedia version 5.2,38 and LncRRIsearch39 servers algorithms.40 In the next stage, we designed a network between selected hub genes and predicted lncRNAs.41
In-silico screening of medicinal chemistry compounds of phytochemicals cocktail
Literature review specified the main bioactive compounds of DA, including coumarin, gallic acid, linalool, α-curcumene, spathulenol, α-eudesmol, anthocyanin (flavylium), selinene, curzerene, coumaric acid, caffeic acid, chlorogenic acid, phytol, hexadecanoic acid, caryophyllene, 3,7,11,15-tetramethyl-2-hexadecen-1-ol, curcumene, Ylangene, gurjunene, barbatene, isohibaene. These bioactive compounds with respective doses in the D. aucheri’s extract were mentioned in Table 1.
Table 1.
Molecular docking of bioactive compounds presented in phytochemicals cocktail
|
No.
|
Phytochemical Compound Name
|
PubChem ID
|
Volume (mg/g or %)
|
Molecular docking: binding affinity (kcal/mol) in RMSD < 2
|
|
CTNNB1
|
SIRT1
|
| 1 |
Coumarin |
323 |
49.65% |
-5.4 |
-5.8 |
| 2 |
Gallic acid |
370 |
0.17 mg/g |
-6.0 |
-6.2 |
| 3 |
Linalool |
6549 |
0.37% |
-4.3 |
-5.4 |
| 4 |
α-Curcumene |
92139 |
1.78% |
-5.2 |
-6.7 |
| 5 |
Spathulenol |
92231 |
6.68% |
-6.3 |
-6.4 |
| 6 |
Α-Eudesmol |
92762 |
7.72% |
-6.3 |
-6.4 |
| 7 |
Anthocyanin (Flavylium) |
145858 |
1.49 mg/g |
-6.4 |
-7.8 |
| 8 |
Selinene |
442393 |
2% |
-6.1 |
-6.9 |
| 9 |
Curzerene |
572766 |
18.7% |
-6.0 |
-6.1 |
| 10 |
Coumaric acid |
637542 |
0.528 mg/g |
-5.3 |
-6.1 |
| 11 |
Caffeic acid |
689043 |
1.65 mg/g |
-5.6 |
-6.5 |
| 12 |
Chlorogenic acid |
1794427 |
1.14 mg/g |
-6.4 |
-8.1 |
| 13 |
Phytol |
5280435 |
14.92% |
-4.4 |
-6.0 |
| 14 |
Hexadecanoic acid |
5282743 |
8.09% |
-4.0 |
-5.2 |
| 15 |
Caryophyllene |
5354499 |
31.29% |
-5.8 |
-6.4 |
| 16 |
3,7,11,15-tetramethyl-2-hexadecen-1-ol |
5366244 |
8.70% |
-4.2 |
-5.8 |
| 17 |
Curcumene |
6428461 |
0.47% |
-5.1 |
-7.4 |
| 18 |
Ylangene |
6432119 |
0.49% |
-6.2 |
-6.2 |
| 19 |
Gurjunene |
6450812 |
9.84% |
-6.0 |
-6.3 |
| 20 |
Barbatene |
14109421 |
0.69% |
-6.1 |
-6.1 |
| 21 |
Isohibaene |
59284349 |
6.16% |
-6.8 |
-7.7 |
We specified that potential cut points in the PPIs network are CTNNB1 and SIRT1, with the highest degree and the most betweenness centrality. CTNNB1 and SIRT1 might be drugable proteins as a new therapeutic approach for hypothyroidism and metabolic bone diseases based on enrichment outputs and browsing molecular signaling pathways in the DAVID database.42 Thus, the Protein Data Bank database (PDB) 49 selected these proteins' best X-ray crystallography. We used Chimera 1.8.1 software to prepare and optimize protein structure.
Based on the literature review and brows of evidence, we found that the phytochemicals cocktail is an effective medicinal herbal as an anti-diabetic, natural cholesterol reducer, and regulator of thyroid hormones.16 Hence, we reviewed the evidence to recognize special bioactive compounds in the phytochemicals cocktail. Accordingly, we browsed the PubChem database to obtain these bioactive compounds' three-dimensional (3D) structures in SDF format. In the next stage, we provided a library that included 3D structures of bioactive compounds using Open Bable software for molecular docking techniques. The dimensional space of vina search was defined for CTNNB1 and SIRT1 proteins surface molecular docking in x = 75.0, y = 68.0, z = 132.0 Å, and x = 76.0, y = 65.0, z = 69.0 Å respectively in PyRx software. Vina space search dimensions are a variable parameter based on the volume of the 3D structure of proteins. Based on previous molecular docking studies, the binding affinity of ligand-macromolecule interactions is accepted in the < -5 kcal/mol and RMSD < 2 as efficient computing. Moreover, we approved binding affinities < -5 kcal/mol and RMSD < 2 in this phytochemical analysis for the efficacy computing of bioactive compounds. For the detection of the interaction between the medicinal chemistry compounds of the phytochemicals cocktail and macromolecules (CTNNB1 and SIRT1), we applied PyMOL and BIOVIA Discovery Studio Visualizer software.
Induced hypothyroidism in mice
Based on the literature review, we selected 8 mg/kg propylthiouracil (PTU) as the optimal dose to induce hypothyroidism in mice. In this study, the PTU was administered by intraperitoneal (IP) (8 mg/kg) for 30 days.24
Phytochemicals cocktail extraction and consumption
DA plant extract bioactive compounds were dried in the darkroom and powdered by an electric mixer according to the hydro distillation protocol in previous studies. The plant extraction is performed using the Soxhlet technique. The plant materials (50 g) were extracted using 200 mL of 95% ethanol for 24 hours using the Soxhlet technique. The temperature did not exceed the solvent's boiling point throughout the extraction process. After filtering the extract with Whitman No. 1 filter paper, the extract was concentrated using a rotary evaporator at 40 ºC.
The extraction percentage was approximated to be 100%, indicating that the whole amount of the desired substance was successfully obtained. Furthermore, it was determined that each ml of the extract contained an equivalent of 1 mg of the plant material. Literature review specified the main bioactive compounds of DA, including coumarin, gallic acid, linalool, α-curcumene, spathulenol, α-eudesmol, anthocyanin (flavylium), selinene, curzerene, coumaric acid, caffeic acid, chlorogenic acid, phytol, hexadecanoic acid, caryophyllene, 3,7,11,15-tetramethyl-2-hexadecen-1-ol, curcumene, Ylangene, gurjunene, barbatene, isohibaene. These bioactive compounds with respective doses in the D. aucheri's extract were mentioned in Table 1. We used a 0.4 mg/kg phytochemicals cocktail for two months/5 days in this study.
Aerobic exercise protocol
For eight weeks, hypothyroidism in mice was implemented for AE on the motorized treadmill. The type of AE was moderate to high-intensity interval training. After adaptation for one week, the protocol training's time, intensity, and reputation were gradually enhanced to finally reach ~ 75% VO2 max (45 min, slope 0, 25 m/min). The intensity and speed of the protocol exercise were set to rise at a rate of 3 m/min and 5 % VO2 max, from 10 m/min to 20% VO2 max to reach 25 m/min and ~ 75% VO2 max. Moreover, after the last session of the protocol exercise and consumption of the phytochemicals cocktail, exhaustion tests were conducted on the motorized treadmill.39,43
Hormones measurement
After AE training and Phytochemicals cocktail consumption, the mice were sacrificed, and the serum was separated by centrifuge (3100 g, 5 min, 4 °C) the level concentration of the T3 (MYBIOSOURCE, MBS262762), T4 (MYBIOSOURCE, MBS700665), and TSH (MYBIOSOURCE, MBS455232) were measured to verify hypothyroidism. Moreover, the level concentration of Sclerostin (Abcam, ab213889) and Dickkopf-1 (DKK1) (R&D Systems, DY1765) was measured to assess biomarkers bone. In this study, we collected 1 ml of blood from each mouse. Blood-containing tubes were centrifuged for 5 min at 4000 × g at 4 °C to separate serum. All samples were frozen in liquid nitrogen and stored at -80 °C until further use. Serum T3, T4, and TSH levels were measured using a commercially available enzyme-linked immunosorbent assay ELISA Kit. T3 level with a sensitivity of less than 20 ng/mL, T4 level with a sensitivity of 0.05 ng /mL, and TSH level with a sensitivity of 1.17 pg/mL were measured by ELISA method to confirm hypothyroidism. It is worth mentioning that the coefficient of change in these kits is between 12-15%. In addition, the sensitivity and range of the Sclerostin kit were 10 pg/mL and 15.6 pg/mL-1000 pg/mL, respectively. Moreover, the range of the DKK1 kit was 46.9 - 3,000 pg/mL.
RNA extraction and qPCR-real time
Total RNAs of the bone tissue samples were isolated using Trizol reagent (Thermo Scientific, USA) based on the manufacturer's protocol. The quality of RNA was evaluated with NanoDrop spectrophotometry (Thermo Scientific, Waltham, MA, USA). The cDNA synthesis was conducted based on the manufacturer's protocol (TaKaRa, Kusatsu, Shiga Prefecture, Japan). Primers of mRNA and long non-coding RNA were designed in Oligo 7 and Beacon Designer 7 software. Furthermore, Real-time qPCR was conducted using SYBR Green dye (TaKaRa, Kusatsu, Japan) on a Rotor-Gene 6000 instrument (Corbett Life Science, Mortlake, Australia). This study used 18S ribosomal RNA (18S rRNA) as a reference gene to normalize the expression of genes and long non-coding RNAs (Table 2).
Table 2.
Primer list
|
Gene
|
Forward primer (5`-3`)
|
Reverse primer (5`-3`)
|
Annealing temperature (°C)
|
| Ppar-ɑ |
ACTTGCTCACTACTGTCCTT |
ATGCTGGTATCGGCTCAATA |
60 |
| Ncor1 |
CTGGTCTTTCAGCCACCATT |
CCTTCATTGGATCCTCCATC |
60 |
| Ctnnb1 |
AGAACACTAATTCATAATCACGCT |
GGCTCAAATAACACCTCTTACTG |
60 |
| Adipoq |
AGGCCGTTCTCTTCACCTACG |
GACTTGGTCTCCCACCTCCAGA |
60 |
|
Sirt1
|
GGCAGTAACAGTGACAGT |
CTCTCCGTATCATCTTCCAA |
61 |
| HOTAIR |
TTGCTCTCCTTATCATCC |
ATTAGTGCCTCTCAGTCC |
51 |
| TUG1 |
CTCTGGAGGTGGACGTTTTGT |
GTGAGGTCGTGTCTCTCTTTTCTC |
51 |
| H19 |
GGAATGTTGAAGGACTGAGGG |
GTAACCGGGATGAATGTCTGG |
60 |
| MEG3 |
ACAAGCGACAAAGAGGAT |
ATGAACACGAGCACAGAT |
53 |
|
18s rRNA
|
CGGACACGGACAGGATTG |
TCGCTCCACCAACTAAGAAC |
59 |
Pathological analysis
After the last treatment, the mice were sacrificed, and the femur bone was separated for histological and pathological analysis. This study fixed the samples in 10% buffered formalin decalcified using an EDTA and HCl (Richard-Allan Scientific, Kalamazoo, MI, USA). After That, the bone samples were stained with hematoxylin and eosin (H&E), and Mason's Trichrome.
Biomechanical assessment
The biomechanical parameters of Tibias from the respective groups were analyzed. The bones were subjected to a three-point bending test using a Zwick/Roell Group material testing apparatus (model Z 2.5 H 15WN, UIm, Germany) until the bone was fractured. All of the bones were positioned in a consistent orientation within the testing apparatus. In this experiment, two loading points were utilized, positioned at a distance of 13 mm from each other, to mount each bone. Following this, a press head was engaged to apply pressure to the central region of the bone shafts until fracture took place. The material testing instrument took data from the load-deformation curve and automatically recorded it. The parameters were calculated, including maximum force (N), stress at high load (N/mm2), energy absorption (N/mm), and bending stiffness (N/mm).
Statistical analysis
The analysis variances were estimated by GraphPad Prism Software (Version 9, GraphPad Software Inc., La Jolla, CA). To ensure the normal distribution, the Shapiro-Wilk test was used. Based on the results, the P value based on the Shapiro-Wilk test was more than 0.05. Hence, the parametric test was employed to analyze the data. Moreover, all data were computed by one-way analysis of variance (ANOVA) with Tukey's post hoc test. Results are shown as mean ± standard deviation (SD), and differences at P < 0.05 indicated the significance in all analyses.
Results
Identification of hub genes, regulatory factors, and molecular signaling pathways based on computational molecular biology
Based on R statistical programming software output of analysis metabolic bone diseases microarray dataset, among 22284 genes, we found 1282 genes with significant differential expression considering P< 0.05 between low bone density and high bone density in premenopausal conditions. Hence, we have shown considerable differential gene expression with P< 0.05 in the heatmap diagram (Fig. 2A). Moreover, we classified significant genes based on logFC + 0.2 and detected 647 genes with upregulation patterns and 636 genes with downregulation patterns. Calculation of visualizing parameters of protein-protein interactions network of complete differential genes expression list highlighted 109 genes as hub genes in this network. We also constructed a PPIs network with network diameters and modularity using Gephi software (Fig. 2B).
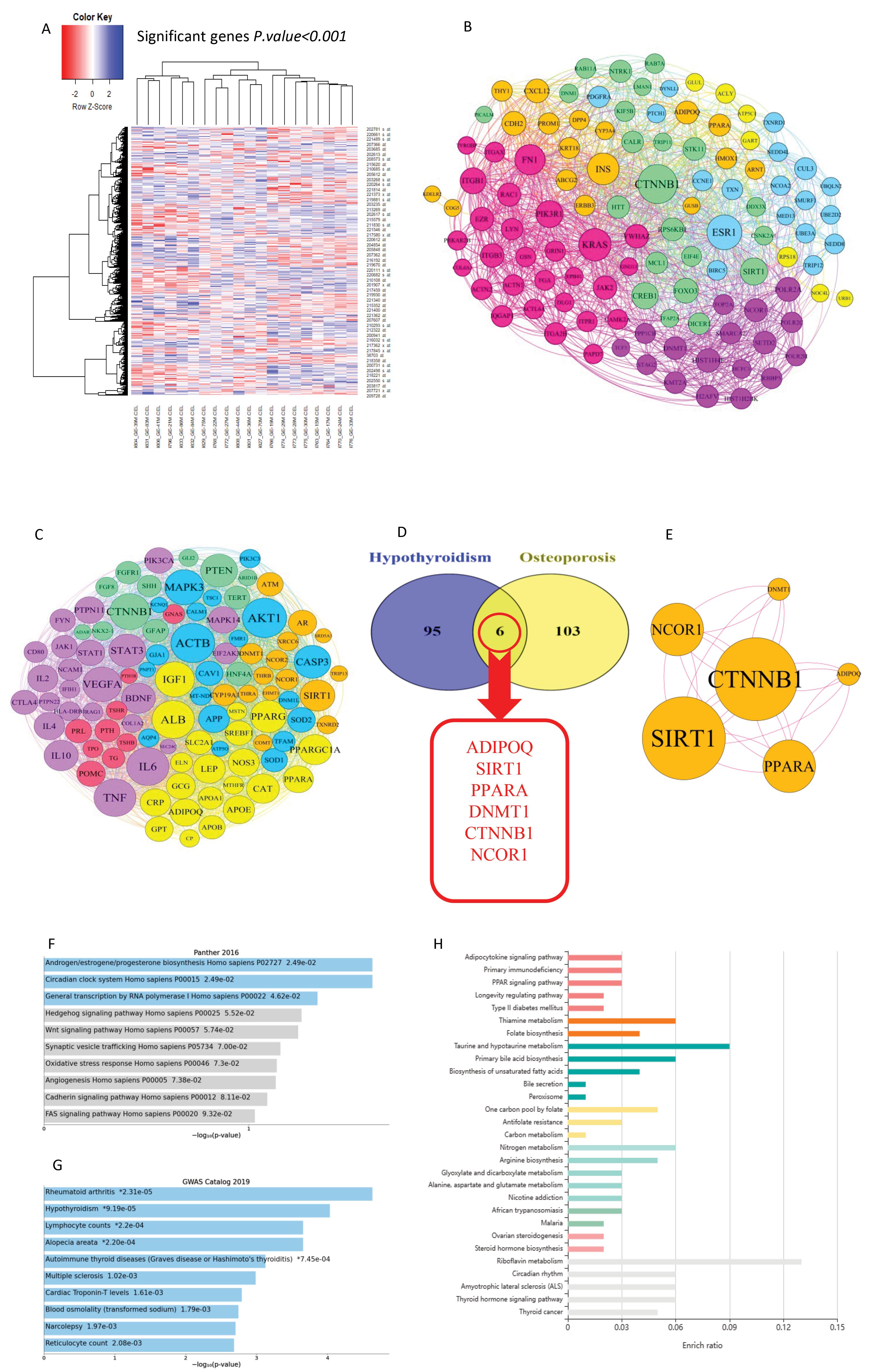
Fig. 2.
Bioinformatics analysis. A: The heatmap of significant genes with differential expression comparison between low bone density and high bone density showed that 807 genes downregulated and 552 genes overexpressed. B: A protein-protein interactions network of hub genes in bone metabolic disorder considering the modularity of genes in the functional mechanisms. C: A network of protein-protein interactions was constructed, including hub genes involved in hypothyroidism, by considering the modularity of genes involved in functional pathways. D: Common hub genes between hypothyroidism and bone metabolism disorder containing ADIPOQ, SIRT1, PPARA, DNMT1, NCOR1, CTNNB1. E: Network construction of common hub genes in hypothyroidism and bone metabolism disorder to highlight the chief cutpoints with the highest degree and the most betweenness centrality. F-H: Browsing of common hub genes in enrichment database to mark crucial molecular signaling pathways involved in bone metabolism pathogenesis manifested that adipocytokine activation signaling, oxidative stress, peroxisome, PPARs pathways, Wnt-B catenin signaling pathway, primary immune deficiency, and metabolism were vital molecular signaling pathways involved in imbalance osteoblast/osteoclast.
.
Bioinformatics analysis. A: The heatmap of significant genes with differential expression comparison between low bone density and high bone density showed that 807 genes downregulated and 552 genes overexpressed. B: A protein-protein interactions network of hub genes in bone metabolic disorder considering the modularity of genes in the functional mechanisms. C: A network of protein-protein interactions was constructed, including hub genes involved in hypothyroidism, by considering the modularity of genes involved in functional pathways. D: Common hub genes between hypothyroidism and bone metabolism disorder containing ADIPOQ, SIRT1, PPARA, DNMT1, NCOR1, CTNNB1. E: Network construction of common hub genes in hypothyroidism and bone metabolism disorder to highlight the chief cutpoints with the highest degree and the most betweenness centrality. F-H: Browsing of common hub genes in enrichment database to mark crucial molecular signaling pathways involved in bone metabolism pathogenesis manifested that adipocytokine activation signaling, oxidative stress, peroxisome, PPARs pathways, Wnt-B catenin signaling pathway, primary immune deficiency, and metabolism were vital molecular signaling pathways involved in imbalance osteoblast/osteoclast.
On the other hand, an analysis of gene lists associated with hypothyroidism was based on visualizing network parameters, which specified 100 genes as hub genes in the pathogenesis of hypothyroidism. We also designed a PPI network for hypothyroidism hub genes with network diameters and modularity in Gephi software (Fig. 2C). We utilized the VENN diagram tool to determine common hub genes between hypothyroidism and metabolic bone diseases and showed that ADIPOQ, SIRT1, PPARA, DNMT1, NCOR1 CTNNB1 have collaborated in hypothyroidism and metabolic bone pathogenesis (Fig. 2D and 2E).
Enrichment of common genes between hypothyroidism genes list and metabolic bone diseases marked that adipocytokine, oxidative stress, peroxisome, PPAR, Wnt-B catenin, primary immune deficiency, and metabolism are critical molecular signaling pathways associated with these hub genes (Fig. 2 F-H). Based on enrichment outputs and browsing molecular signaling pathways in the DAVID server, we found that CTNNB1 and SIRT1, with the highest degree and the most betweenness centrality in the PPIs network, are potential cut points and might be druggable proteins as the new therapeutic approach in hypothyroidism and metabolic bone diseases.
lncRNAs in metabolic bone diseases related to hypothyroidism status
Based on R statistical programming analysis of GSE56815 data set for specified variable transcriptome expression in bone metabolism, considering low-bone density and high bone density in premenopausal women status.44 We discovered CTC-338M12.4, LINC00474, LINC02809, RPARP-AS1, HGC6.3, LINC00965, ZNF217-AS1, LOC100505498, and DDR1DT as potential non-coding RNAs in the bone metabolism. However, the main role of these markers in cellular, molecular, and signaling functions is unclear, and their effect on bone metabolism in the hypothyroidism-bone axis is still unknown. On the other hand, browsing lncRNAs prediction databases based on genes, disease, and signaling pathways led to the identification of TUG1, HOTAIR, MEG3, and H19 as regulation factors in the biological processes of metabolic bone pathogenesis. TUG1 could regulate the CTNNB1 and SIRT1 gene expression, affecting the Wnt/B catenin signaling pathway. On the other hand, we found that MEG3 and H19 could be targeted and regulated by DNMT1. Also, we recognized HOTAIR as a regulatory factor with roles in histone modification, sponge, and interaction with the protein that influenced the Wnt/B-catenin signaling pathway and regulated osteogenic differentiation and bone formation. Moreover, we found that HOTAIR could target DNMT1 and regulate apoptosis processes. A network between selected hub genes and predicted lncRNAs was designed, as shown in Fig. 3.
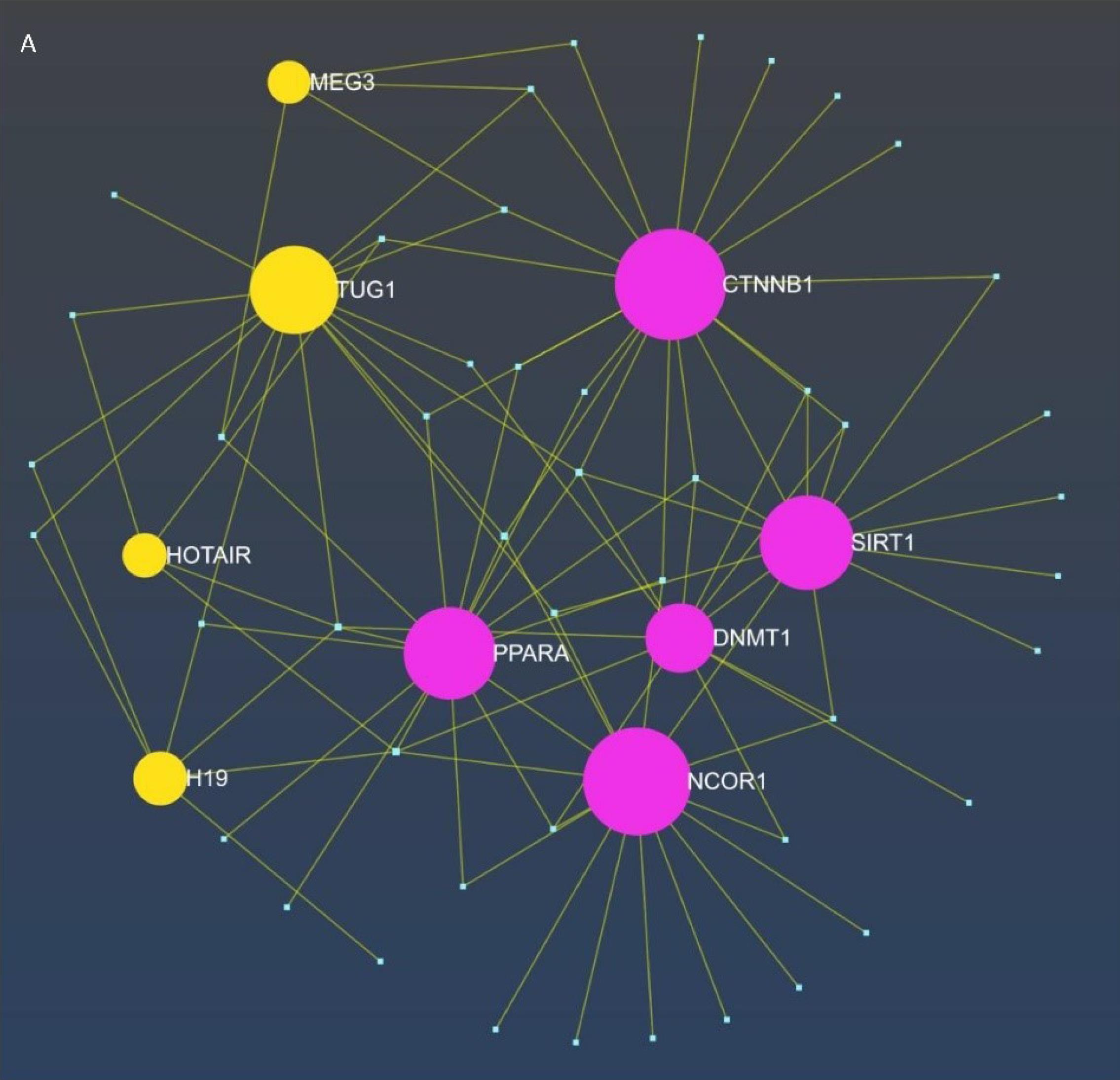
Fig. 3.
The display of interaction between hub genes, microRNA and predicted lncRNAs as ceRNA network. TUG1 could regulate the expression of the CTNNB1 and SIRT1 genes, hence influencing the Wnt/β-catenin signaling pathway. Conversely, we discovered that MEG3 and H19 may be targeted and controlled by DNMT1. We identified HOTAIR as a regulatory factor involved in histone modification, acting as a sponge, and interacting with proteins that affect the Wnt/β-catenin signaling cascade, hence regulating osteogenic differentiation and bone formation. Furthermore, we discovered that HOTAIR can target DNMT1 and modulate apoptotic processes.
.
The display of interaction between hub genes, microRNA and predicted lncRNAs as ceRNA network. TUG1 could regulate the expression of the CTNNB1 and SIRT1 genes, hence influencing the Wnt/β-catenin signaling pathway. Conversely, we discovered that MEG3 and H19 may be targeted and controlled by DNMT1. We identified HOTAIR as a regulatory factor involved in histone modification, acting as a sponge, and interacting with proteins that affect the Wnt/β-catenin signaling cascade, hence regulating osteogenic differentiation and bone formation. Furthermore, we discovered that HOTAIR can target DNMT1 and modulate apoptotic processes.
Drug affinity outcomes based on pharmacophore modeling and molecular docking
Based on In-silico screening and biomedicinal chemistry analysis, we estimated the binding affinity of bioactive compounds in the Phytochemicals cocktail on CTNNB1 and SIRT1 as the hub proteins with the ability to cut points involved in the Wnt/B-catenin signaling pathway and metabolism of bone tissue (Table 1).
In this in silico study, based on the evidence, we found 21 effective bioactive compounds including coumarin, gallic acid, linalool, curcumene, spathulenol, alpha-eudesmol, flavylium, beta-selinene, curzerene, coumaric acid, caffeic acid, chlorogenic acid, phytol, hexadecenoic acid, caryophyllene, phytol (3,7,11,15-tetramethyl-2-hexadecen-1-OL), curcumin, Ylangene, gurjunene, barbatene, and isohibaene in DA extract. Molecular docking computing estimated that Isohibaene binds to the B-catenin protein with the highest binding affinity score (-6.8 kcal/mol) among the bioactive compounds library (Fig. 4A). On the other hand, we indicated that Chlorogenic Acid could be bound to the active site of the SIRT1 protein surface with the most binding affinity score (-8.1kcal/mol) (Fig. 4B).
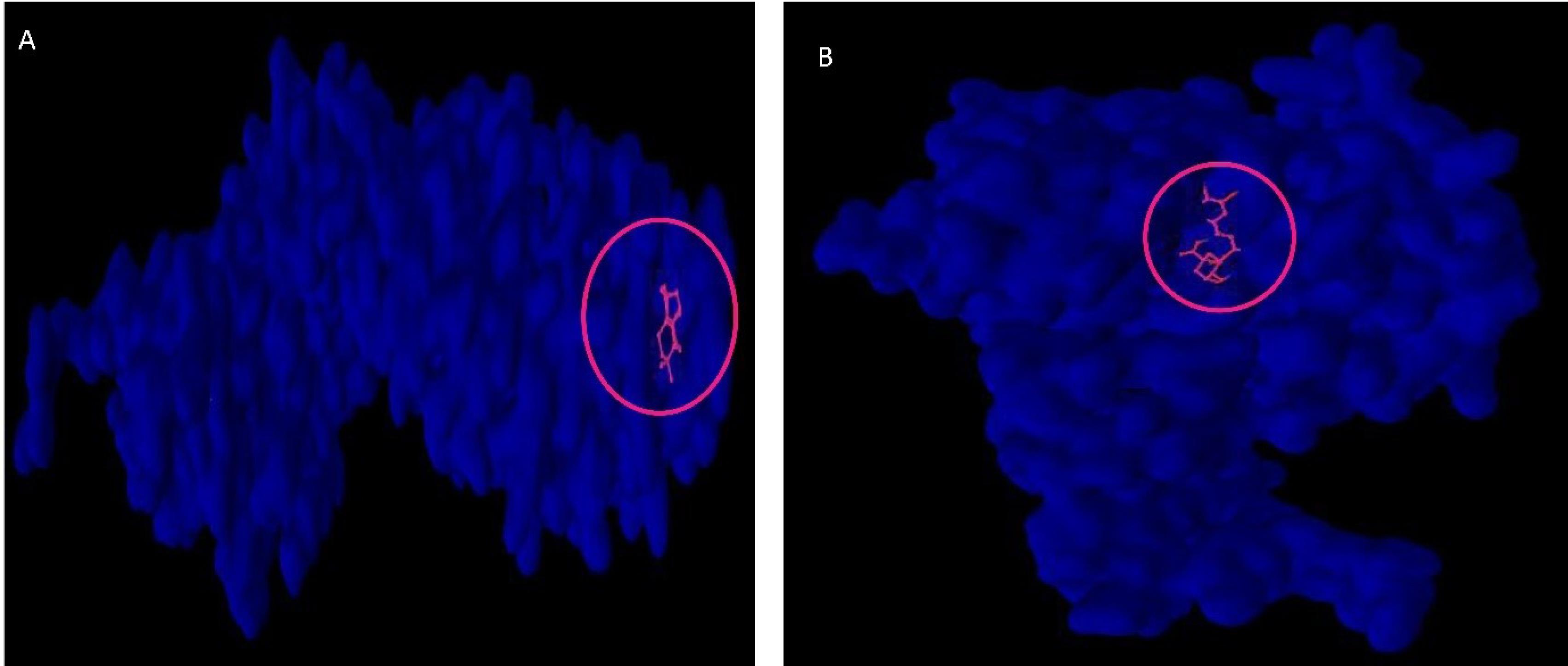
Fig. 4.
Phytochemoinformatic analysis to determine the effectiveness and safety of phytochemicals cocktail bioactive compounds in bone metabolism disorder induced by hypothyroidism. A: Molecular docking computing estimated that Isohibaene with the highest binding affinity score (-6.8 kcal/mol) among 21 bioactive compounds of phytochemicals cocktail bind to the B-catenin protein. B: Among the 21 bioactive compounds presented in phytochemicals cocktail, chlorogenic acid has the highest binding affinity (-8.1 kcal/mol) to the active site of the SIRT1 protein.
.
Phytochemoinformatic analysis to determine the effectiveness and safety of phytochemicals cocktail bioactive compounds in bone metabolism disorder induced by hypothyroidism. A: Molecular docking computing estimated that Isohibaene with the highest binding affinity score (-6.8 kcal/mol) among 21 bioactive compounds of phytochemicals cocktail bind to the B-catenin protein. B: Among the 21 bioactive compounds presented in phytochemicals cocktail, chlorogenic acid has the highest binding affinity (-8.1 kcal/mol) to the active site of the SIRT1 protein.
Based on the literature review, the molecular and cellular signaling pathways regulated by bioactive compounds are presented in Table 1. Data mining about Chlorogenic acid compound effects has shown that this phytochemical formula is pivotal as the potential antioxidant in scavenging free radicals (Table 3). It is worth noting that the total bioactive compounds in this computational chemistry analysis bonded to the SIRT1 protein with an acceptable binding affinity score ( < -2 kcal/mol). Based on the cheminformatic analysis, the interaction between medicinal chemistry compounds of phytochemicals cocktail and macromolecules (SIRT1 and CTNNB1) were visualized in Fig. 5.
Table 3.
Molecular and cellular signaling pathways that are triggered/suppressed by bioactive compounds based on the literature review
|
Bioactive compounds
|
Molecular and cellular signaling pathways
|
Ref.
|
| Coumarin |
-
Interfere with bone metabolism.
-
Inhibit osteoclast function.
-
Downregulate RANKL-RANK signaling, NF-κB, MAPK, Akt, Ca2+ signaling, nuclear factor of activated T-cells, cathepsin K, TRAP, and MMP-9 for osteoclast development.
-
Improving bone health
|
45
|
| Gallic acid |
-
Inhibited RANKL
-
Blocking Akt, ERK, and JNK pathways and suppressed osteoclastogenesis-related marker expression, including a nuclear factor of the activated T-cell cytoplasmic 1 (NFATc1), c-Fos, and cathepsin K
-
Reduces F-actin formation and osteoclast resorptive activity
-
Preventing bone loss
|
46
|
| Linalool |
-
Cytotoxic activity
-
Antiproliferative
-
Antioxidant
|
47
|
| α-Curcumene |
-
Antimicrobial
-
Larvicidal
-
Pheromone
-
Antioxidant
-
Hepatoprotective
-
COX2 inhibitor
-
Antiandrogenic
-
Analgesic
|
| Spathulenol |
-
Regulating TGF-β1/Smads and PI3K/Akt pathways
-
Inhibiting hepatic stellate cell activation
-
Reducing extracellular matrix secretion and deposition
|
48
|
| α-Eudesmol |
-
Preventing the increase in gene expression of fibrosis markers procollagen I and procollagen III
-
Alleviating the collagen volume fraction
-
Modulation of JAK/STAT, NF-κB, ERK1/2, MAPK, and CREB signaling pathways
|
1
|
| Anthocyanin |
-
C-Fos, NF-κB, JNK, Ca2 + , ROS, P38, ERK1/2, JNK, and MAPK signaling pathways are essential in RANK signal-mediated osteoclast generation. The nuclear factor of activated T-cells 1(NFATc1) is a major transcription factor and an essential target gene of most pathways regulating osteoclastic differentiation. Anthocyanins suppress osteoclast formation and differentiation through more than one of these pathways.
|
49
|
| Coumaric acid |
-
Activated the peripheral AMPK pathway
-
Enhanced hypothalamic leptin sensitivity
-
Improved blood glucose control, glucose tolerance, and insulin sensitivity
-
increases chondrocyte proliferation and/or affects the expression of IGF-1
-
Antioxidant, anti-inflammatory, anti-diabetic, anti-ulcer, anti-platelet, anti-cancer activities
-
The expression of main transcriptional factors such as CCAAT/enhancer binding protein-alpha (C/EBP-α) and PPAR-γ2 decreased.
-
Decreased the expression of SREBP-1, fatty acid synthase, acetyl CoA carboxylase, and adiponectin.
|
50,51
|
| Caffeic acid |
-
Reduced osteoclastogenesis, inhibited osteoblast apoptosis and bone resorption
-
Antioxidant potential
-
Increased expression of osteoblast markers
-
activating NRF2/HO-1 and inhibiting the NF-κB signaling pathway
-
Impact on oxidative stress and RANKL/OPG signals
|
52
|
| Chlorogenic acid |
-
Prevents Osteoporosis by Shp2/PI3K/Akt Pathway
-
Inhibited the phosphorylation of p38, Akt, extracellular signal-regulated kinase (ERK), and inhibitor of nuclear factor-kappa B (IκB), and IκB degradation by RANKL treatment
-
Suppressed the mRNA expression of nuclear factor of activated T cells c1 (NFATc1), TRAP, and OSCAR in RANKL-treated bone marrow macrophages (BMMs)
-
Inhibits Osteoclast Differentiation and Bone Resorption
-
Activating the Nrf2/HO-1 pathway.
-
Activating p21Waf1/Cip1
|
53-55
|
| Phytol |
-
Produces Antihyperalgesic, Anti-inflammatory, and Antiarthritic Effects
-
Suppresses Osteoclast Differentiation and Oxidative Stress through Nrf2/HO-1 Regulation
-
Possible NF-κB Pathway Involvement and Reduced Levels of the Proinflammatory Cytokines TNF-α and IL-6
|
56,57
|
| Caryophyllene |
-
Selective agonist of the cannabinoid receptor type 2 and exerts cannabimimetic anti-inflammatory effects
-
Reduces acute and chronic pain associated with inflammation
-
Reduced TNF-α and interleukin (IL)-1β production
-
Regulates the differentiation of bone marrow cells associated with adipogenesis and osteoblastogenesis
-
Enhance osteoblastogenesis
-
Suppress adipogenesis and osteoclastogenesis
|
58
|
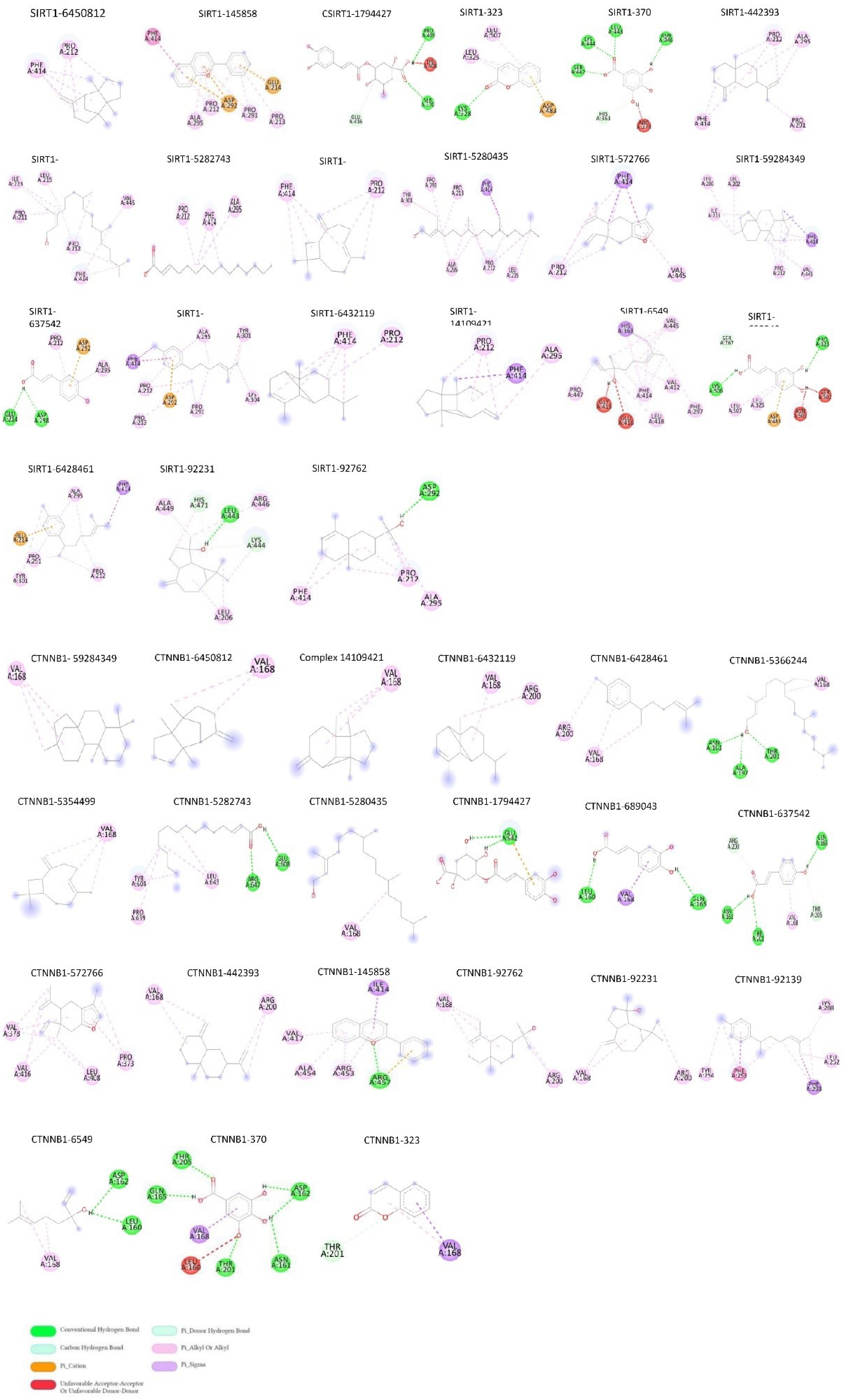
Fig. 5.
Interaction (residual site) between medicinal chemistry compounds of phytochemicals cocktail (based on PubChem ID) and macromolecules SIRT1 and CTNNB1. Different colors in the guidance characterize bond types.
.
Interaction (residual site) between medicinal chemistry compounds of phytochemicals cocktail (based on PubChem ID) and macromolecules SIRT1 and CTNNB1. Different colors in the guidance characterize bond types.
Clinical signs and mortality
A few nonspecific clinical signs were observed in PTU-treated mice during the treatment. During inducing hypothyroidism with PTU, we found decreased skin turgor, rough hair coat, hunched posture, and/or decreased activity. In this study, we did not have morbidity associated with this experiment.
Aerobic exercise and phytochemicals cocktail reversed phenotype feature in hypothyroid mice
This study indicated that calorie intake, lean mass, fat mass, and body weight decreased in hypothyroid mice (Fig. 6a-d). Moreover, phytochemicals cocktail and AE improved calorie intake, lean mass, fat mass, and body weight (Fig. 6a-d). In addition, we found that the T3, T4, and TSH hormone concentrations significantly changed in hypothyroid mice compared with the Normal group (Fig. 6e-g). Furthermore, HYPO-DA and HYPO-AE ameliorated the T3, T4, and TSH hormone concentrations (Fig. 6a-g). Interestingly, calorie intake, lean mass, fat mass, and body weight were enhanced in the HYPO-DA + AE group compared with other groups as well as the level concentration of T3 and T4 was significantly enhanced, and TSH decreased in the HYPO-DA + AE group (Fig. 6a-g).
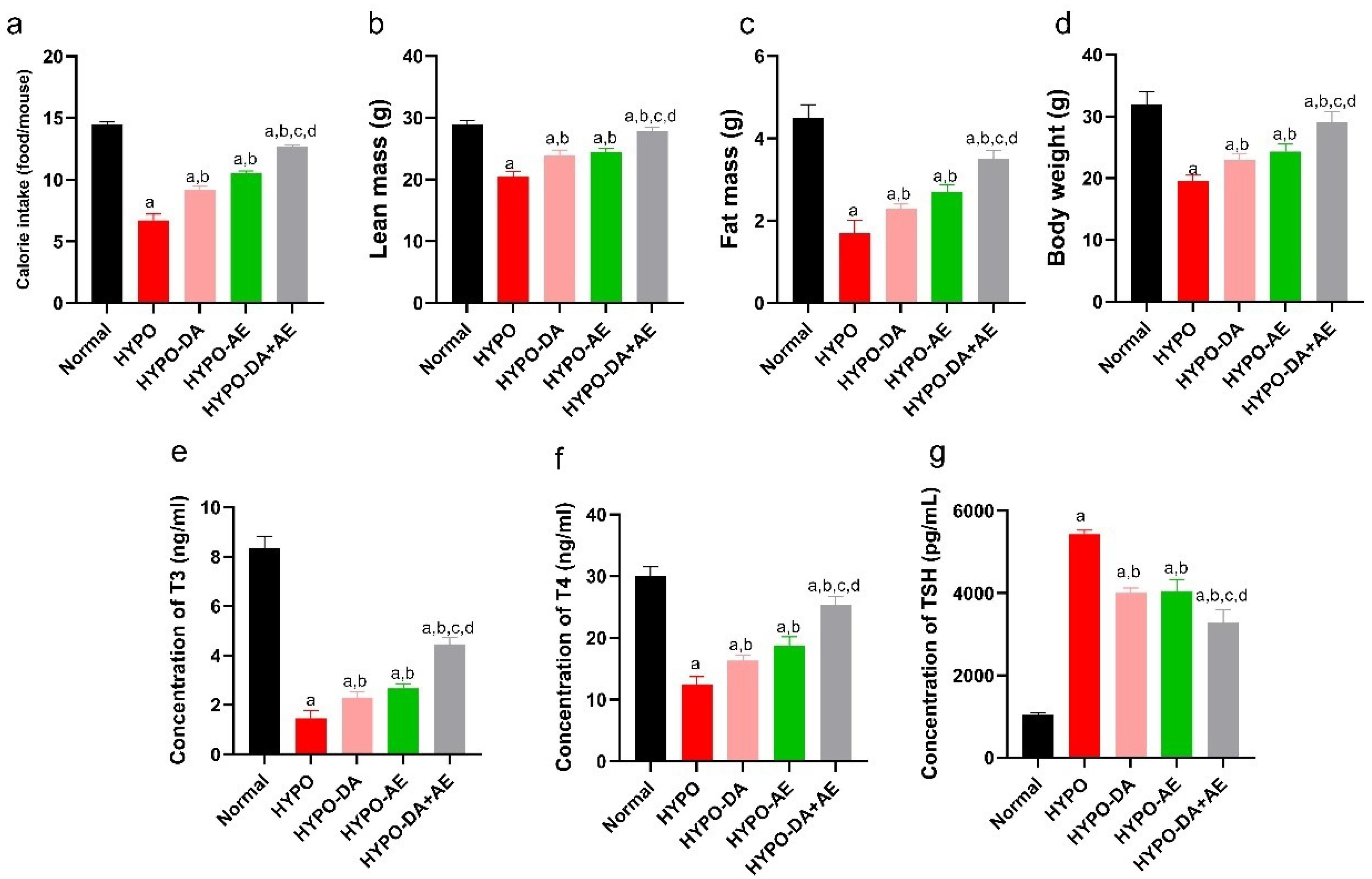
Fig. 6.
Aerobic exercise and the phytochemical cocktail induced alterations in both phenotype and biochemical markers in hypothyroid mice. a: Daily caloric intake for each groups was calculated, (mean ± SD; n = 6, P < 0.05)., b: Lean mass was measured for each groups, (mean ± SD; n = 6, P < 0.05)., c: Fat mass was weighted each groups, (mean ± SD; n = 6, P < 0.05)., d: Body weight was monitored on a weekly, (mean ± SD; n = 6, P < 0.05)., e-g: Serum levels of T3, T4, and TSH were assessed, (mean ± SD; n = 6, P < 0.05). a: shows statistically significant difference with Normal group, b: shows statistically significant difference with HYPO group, c: shows statistically significant difference with HYPO-DA group. d: shows statistically significant difference with HYPO-AE group.
.
Aerobic exercise and the phytochemical cocktail induced alterations in both phenotype and biochemical markers in hypothyroid mice. a: Daily caloric intake for each groups was calculated, (mean ± SD; n = 6, P < 0.05)., b: Lean mass was measured for each groups, (mean ± SD; n = 6, P < 0.05)., c: Fat mass was weighted each groups, (mean ± SD; n = 6, P < 0.05)., d: Body weight was monitored on a weekly, (mean ± SD; n = 6, P < 0.05)., e-g: Serum levels of T3, T4, and TSH were assessed, (mean ± SD; n = 6, P < 0.05). a: shows statistically significant difference with Normal group, b: shows statistically significant difference with HYPO group, c: shows statistically significant difference with HYPO-DA group. d: shows statistically significant difference with HYPO-AE group.
Aerobic exercise and phytochemicals cocktail improved bone remodeling dysregulated in hypothyroid mice
To evaluate bone remodeling, the relative expression of the ALP, Osterix (Osx), Sost, BSP/Collagen1/RANKL, and (Trap) and also the concentration of the Sclerostin and Dkk1 were measured (Fig. 7a-f). Results indicated that the ALP and Osx expression level was reduced in hypothyroid mice vs. the Normal group (Fig. 7a, b). Furthermore, relative expression genes of ALP and Osx were improved in the HYPO-DA and HYPO-AE groups compared with the HYPO group (Fig. 7a, b). Also, HYPO-DA + AE enhanced the expression level of ALP and Osx in comparison with HYPO-DA, HYPO-AE, and HYPO groups (Fig. 7a, b).
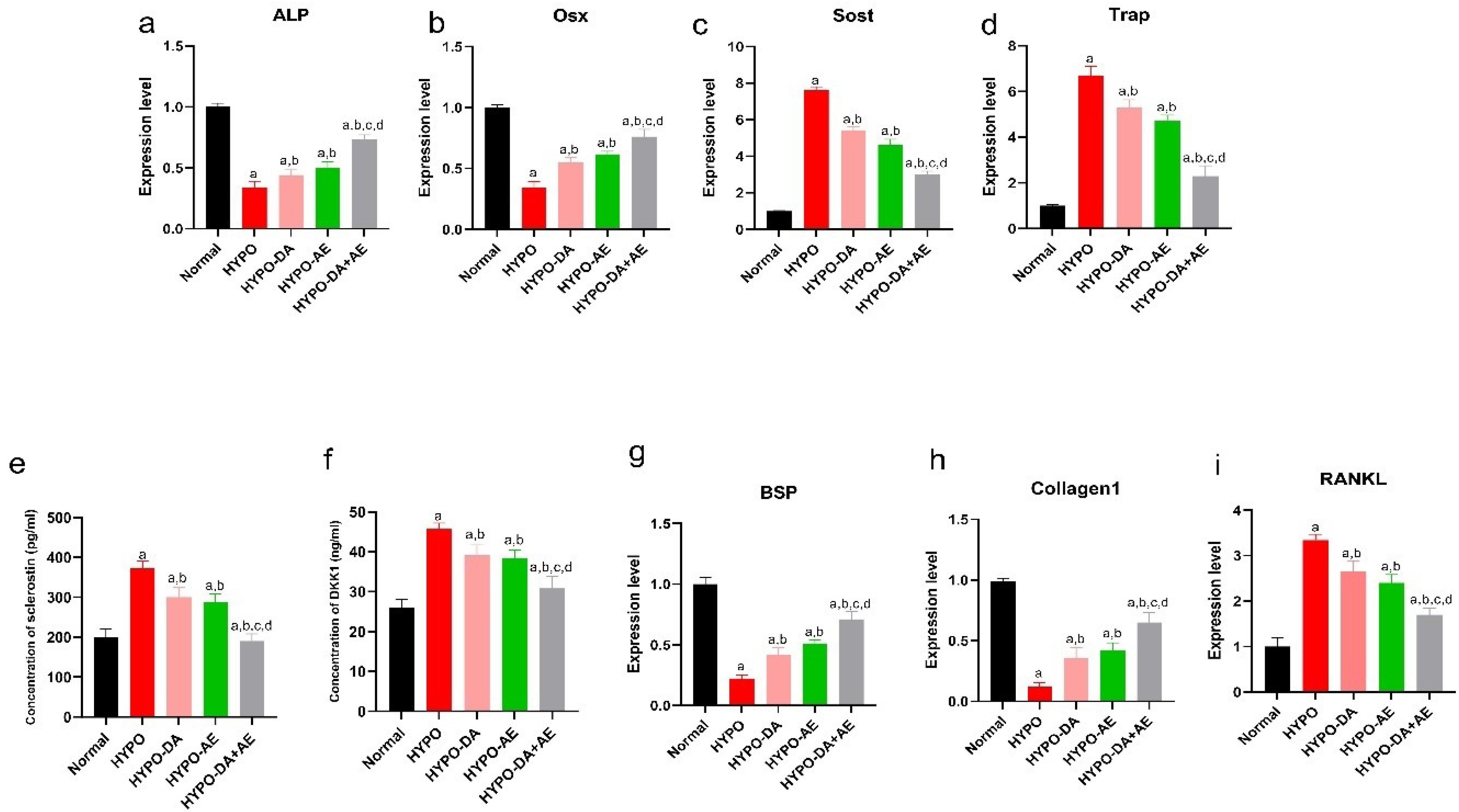
Fig. 7.
Aerobic exercise and the phytochemical cocktail effectively ameliorated the dysregulation of bone remodeling in hypothyroid mice. a-d: The relative expression of the Alp, Osx, Sost, and Trap mRNAs was measured by qPCR Real-Time, (mean ± SD; n = 6, P < 0.05)., e and f: The concentration level of the sclerostin and Dkk1 was assessed by ELISA method for each groups, (mean ± SD; n = 6, P < 0.05)., g-i: The relative expression of the BSP, Collagen1, and RANKL mRNAs was measured by qPCR Real-Time, (mean ± SD; n = 6, P < 0.05). a: shows statistically significant difference with Normal group, b: shows statistically significant difference with HYPO group, c: shows statistically significant difference with HYPO-DA group. d: shows statistically significant difference with HYPO-AE group.
.
Aerobic exercise and the phytochemical cocktail effectively ameliorated the dysregulation of bone remodeling in hypothyroid mice. a-d: The relative expression of the Alp, Osx, Sost, and Trap mRNAs was measured by qPCR Real-Time, (mean ± SD; n = 6, P < 0.05)., e and f: The concentration level of the sclerostin and Dkk1 was assessed by ELISA method for each groups, (mean ± SD; n = 6, P < 0.05)., g-i: The relative expression of the BSP, Collagen1, and RANKL mRNAs was measured by qPCR Real-Time, (mean ± SD; n = 6, P < 0.05). a: shows statistically significant difference with Normal group, b: shows statistically significant difference with HYPO group, c: shows statistically significant difference with HYPO-DA group. d: shows statistically significant difference with HYPO-AE group.
The expression level of the Sost and Trap genes as bone resorption markers was assessed in this study (Fig. 7c, d). Moreover, the Sost and Trap's relative expression significantly increased in the HYPO group compared with the Normal group (Fig. 7c, d). Furthermore, we found that AE and administration of the Phytochemicals cocktail reduced the relative expression of the Sost and Trap genres (Fig. 7c, d). In addition, AE, along with the administration of a phytochemical cocktail (HYPO-DA + AE), predominantly decreased the expression of Sost and Trap (Fig. 7c, d).
On the other hand, we evaluated Sclerostin and Dkk1 concentrations recognized as bone resorption markers (Fig. 7e, f). We revealed that the concentration of Sclerostin and Dkk1 were amplified in the HYPO group (Fig. 7e, f). Moreover, we found that the concentration of Sclerostin and Dkk1 was improved in the HYPO-DA and HYPO-AE groups compared with the HYPO group (Fig. 7e, f). In addition, HYPO-DA + AE significantly decreased the concentration of Sclerostin and Dkk1 compared with HYPO-DA, HYPO-AE, and HYPO groups (Fig. 7e, f). It should be noted that the relative expression level of the BSP/Collagen1/RANKL was dysregulated in the HYPO group (Fig. 7g-i). Moreover, the BSP/Collagen1/RANKL expression level was ameliorated in the HYPO-DA and HYPO-AE groups compared with the HYPO group (Fig. 7e, f). HYPO-DA + AE improved the relative expression of the BSP/Collagen1/RANKL (Fig. 7e, f).
Aerobic exercise and phytochemicals cocktail regulated the Metabolism, Oxidative stress, and Wnt signaling pathway
We revealed that six genes were common between metabolic bone disease and hypothyroidism condition based on artificial intelligence. Furthermore, we found that Adipoq, Dnmt1, Sirt-1, Ppar-ɑ, Ncor1, and Ctnnb1 dysregulated in hypothyroidism conditions (Fig. 8a-f). Our data indicated that the Adipoq and Dnmt1 genes' relative expression decreased, and Sirt-1, Ppar-ɑ, Ncor1, and Ctnnb1 increased in the HYPO compared with the Normal group (Fig. 8a-f). In addition, we demonstrated that AE and administration of 0.4 mg/kg Phytochemicals cocktail(HYPO-DA, HYPO-AE groups) could down-regulated the expression level of the Adipoq and Dnmt1 and upregulated the expression level of the Sirt-1, Ppar-ɑ, Ncor1, and Ctnnb1 (Fig. 8a-f). Notably, we indicated a synergistic effect of AE along with a 0.4 mg/kg Phytochemicals cocktail (HYPO-DA + AE) in bone metabolism compared with other groups (Fig. 8a-f). Based on these results, HYPO-DA + AE significantly decreased the Adipoq and Dnmt1 genes and enhanced the Sirt-1, Ppar-ɑ, Ncor1, and Ctnnb1 genes vs. HYPO-DA, HYPO-AE, and HYPO groups (Fig. 8a-f). Hence, AE along with a 0.4 mg/kg Phytochemicals cocktail might modulate Metabolism, Oxidative stress, and Wnt signaling pathway in the bone of hypothyroid mice.
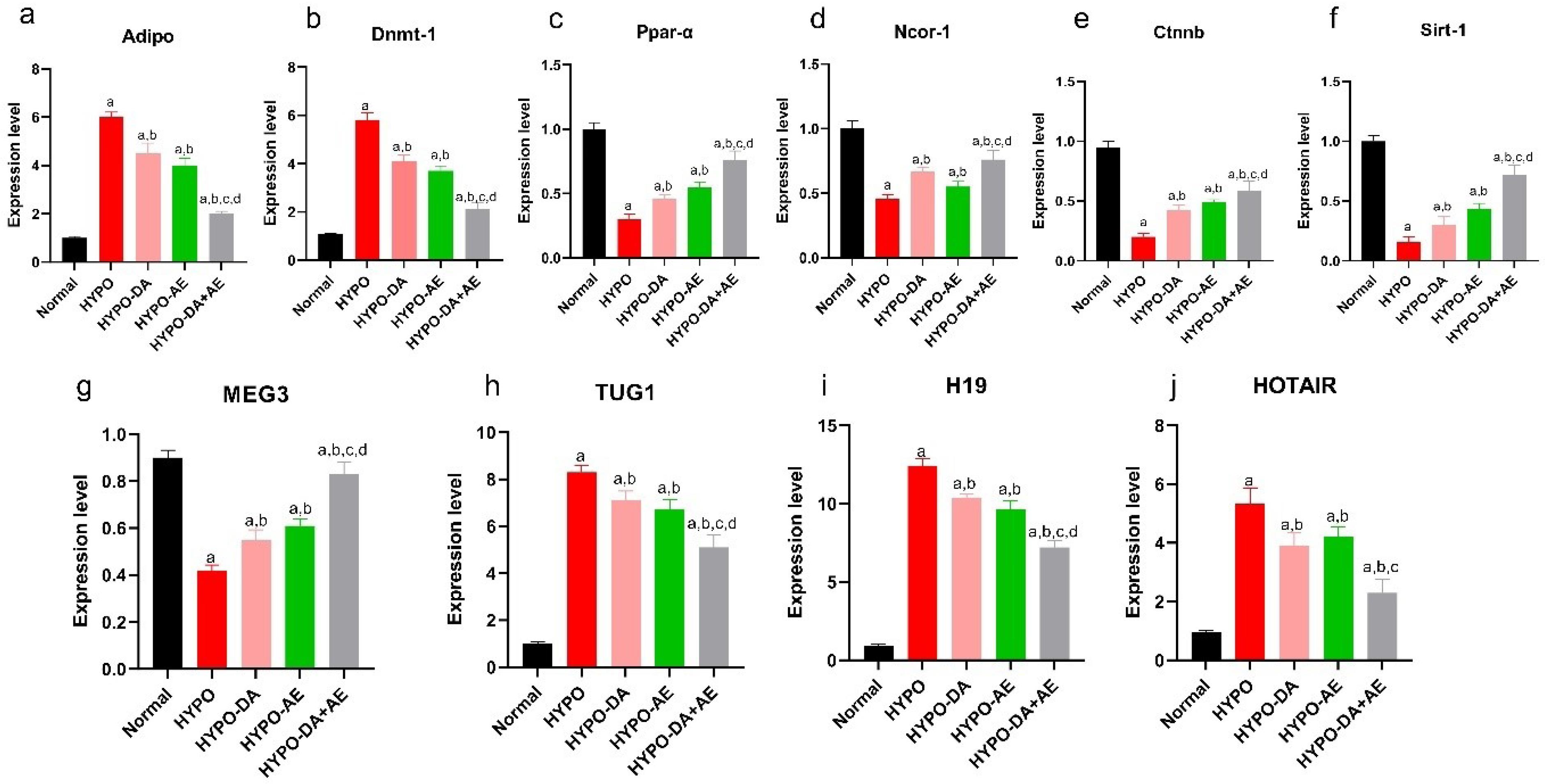
Fig. 8.
Impact of aerobic exercise and phytochemical cocktail on the modulation of mRNA-lncRNA networks in hypothyroid mice. a-f: the relative expression of the Adipoq, Dnmt1, Ppar-ɑ, Ncor1, Ctnnb1, and Sirt-1 in bone tissue, (mean ± SD; n = 6, P < 0.05). g-j: The level expression of the MEG3, TUG1, H19, and HOTAIR, (mean ± SD; n = 6, P < 0.05). a: shows statistically significant difference with Normal group, b: shows statistically significant difference with HYPO group, c: shows statistically significant difference with HYPO-DA group. d: shows statistically significant difference with HYPO-AE group.
.
Impact of aerobic exercise and phytochemical cocktail on the modulation of mRNA-lncRNA networks in hypothyroid mice. a-f: the relative expression of the Adipoq, Dnmt1, Ppar-ɑ, Ncor1, Ctnnb1, and Sirt-1 in bone tissue, (mean ± SD; n = 6, P < 0.05). g-j: The level expression of the MEG3, TUG1, H19, and HOTAIR, (mean ± SD; n = 6, P < 0.05). a: shows statistically significant difference with Normal group, b: shows statistically significant difference with HYPO group, c: shows statistically significant difference with HYPO-DA group. d: shows statistically significant difference with HYPO-AE group.
Aerobic exercise and phytochemicals cocktail modulated mRNA-lncRNAs networks
We assessed the relative expression of the four profiles of lncRNAs (TUG1, HOTAIR, H19, and MEG3), which could bind to the particular genes (Dnmt1, Sirt-1, and Ctnnb1) (Fig. 8g-j). Results indicated that the relative expression of the MEG3 was decreased in the HYPO group compared with the Normal group (Fig. 8g-j). Moreover, we indicated that the TUG1, H19, and HOTAIR expression levels were significantly elevated in the HYPO group compared with the Normal group (Fig. 8g-j). On the other hand, AE and 0.4 mg/kg Phytochemicals cocktail could regulate the lncRNAs expression level, including the MEG3 (down-regulated) and TUG1, H19 HOTAIR (upregulated) compared with the HYPO group (Fig. 8g-j). Notably, the interaction of AE and the Phytochemicals cocktail significantly modulated the MEG3, TUG1, H19, and HOTAIR expression levels. In addition, we revealed that the expression level of the MEG3 was significantly enhanced compared with the other groups (HYPO-DA, HYPO-AE, and HYPO groups) (Fig. 8g-j). Moreover, we have shown that the TUG1, H19, and HOTAIR levels declined compared to HYPO-DA, HYPO-AE, and HYPO groups (Fig. 8g-j).
Aerobic exercise and phytochemicals cocktail changed the biomechanical features
In Table 4, we demonstrated that maximum force (N), stress at high load (N/mm2), energy absorption (N/mm), and bending stiffness (N/mm) were decreased in the HYPO group. In addition, AE and phytochemicals increased the biomechanical features of the HYPO-DA and HYPO-AE groups. Notably, AE along with a phytochemicals cocktail (HYPO-DA + AE group) significantly enhanced the maximum force (N), stress at high load (N/mm2), energy absorption (N/mm), and bending stiffness (N/mm).
Table 4.
Energy absorption, high-stress load, maximum force, and bending stiffness
|
|
Energy absorption
(N/mm)
|
High stress load (N/mm2)
|
Maximum force
(N)
|
Bonding stiffness
(N/mm)
|
P
value
|
| Normal |
39.7 ± 5.1 |
26 ± 1.65 |
45 ± 3.9 |
98.6 ± 6.2 |
< 0.05 |
| HYPO |
13.2 ± 2.52 |
9.2 ± 1.2 |
14 ± 2.1 |
36.3 ± 3.1 |
| HYPO-DA |
25.51 ± 3.78 |
15.1 ± 3.2 |
26 ± 4.6 |
42.7 ± 4.74 |
| HYPO-AE |
27.4 ± 4.23 |
16.8 ± 2.6 |
30 ± 3.7 |
45.3 ± 3.13 |
| HYPO-DA + AE |
32.8 ± 2.69 |
22.3 ± 1.8 |
38 ± 2.8 |
68.21 ± 5.78 |
Osteoclast and osteoblast imbalance in hypothyroid mice
We indicated that AE and a 0.4 mg/kg Phytochemicals cocktail might contribute to the modulation of osteoblast activity and regulate the osteoclast and osteoblast imbalance according to the H&E staining. Furthermore, Masson's trichrome analysis demonstrated that AE and 0.4 mg/kg Phytochemicals cocktail decreased the defect regions vs. the HYPO group (Fig. 9a). Moreover, based on the H&E analysis, we found that the number of osteoclasts and osteoblasts increased in hypothyroid mice compared with the normal group (Fig. 9a). These data were consistent with the increasing concentration of Sclerostin and Dkk1 as well as the relative expression of the Sost and Trap in the HYPO group. In addition, based on Masson's trichrome staining, we revealed that the defect regions in the HYPO group were higher than the Normal group (Fig. 9b).
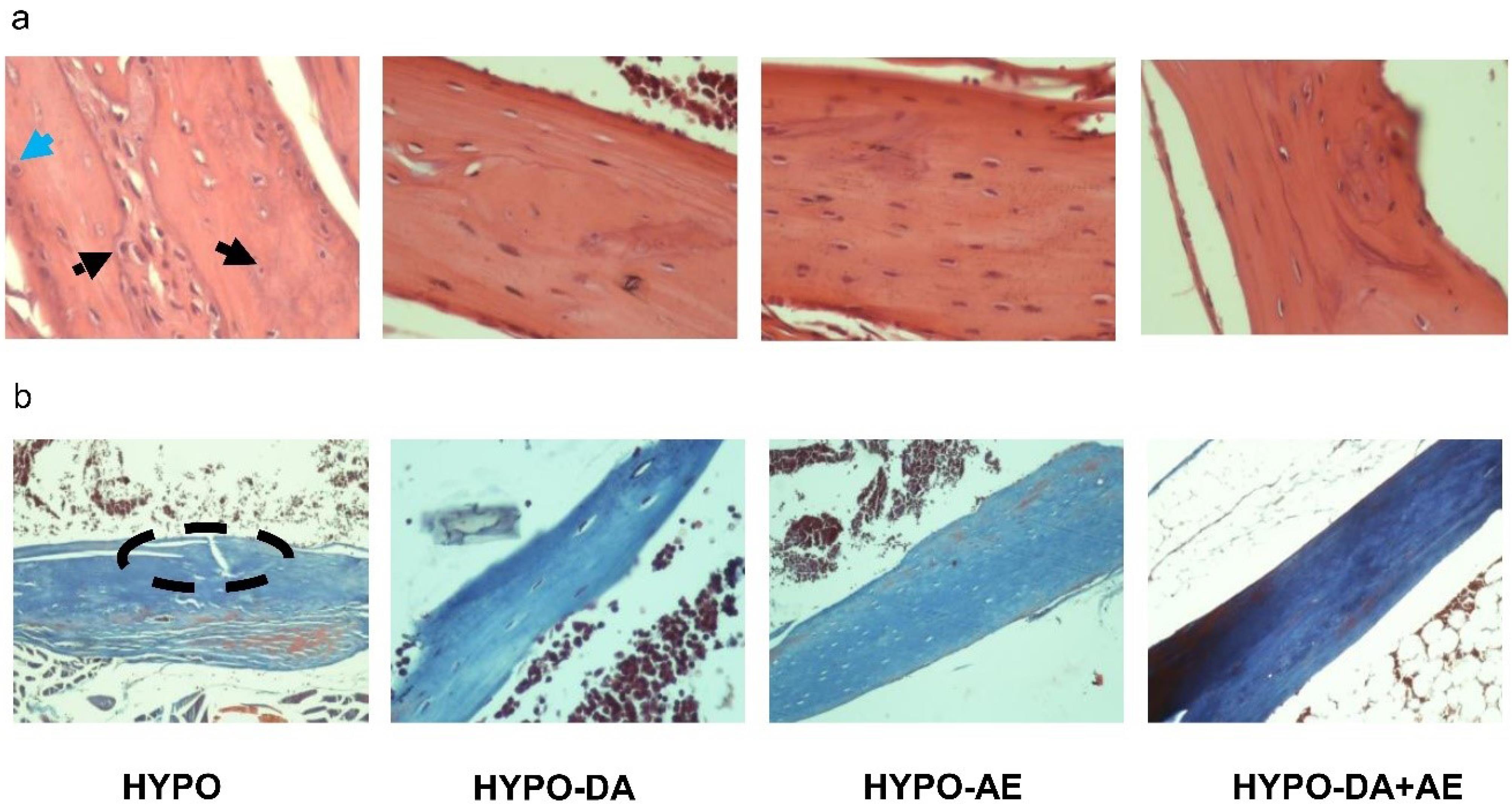
Fig. 9.
Aerobic exercise and phytochemicals cocktail in hypothyroidism mice ameliorated the alternation of histological bone tissue. a and b: H&E and Masson's trichrome staining. The black round dot arrow indicated the osteocytes, the black line arrow indicated the osteoblasts, the blue line arrow indicated the osteoclasts, and the dashed oval indicated the defect bone region.
.
Aerobic exercise and phytochemicals cocktail in hypothyroidism mice ameliorated the alternation of histological bone tissue. a and b: H&E and Masson's trichrome staining. The black round dot arrow indicated the osteocytes, the black line arrow indicated the osteoblasts, the blue line arrow indicated the osteoclasts, and the dashed oval indicated the defect bone region.
Discussion
Hypothyroxinemia in mice could dysregulate bone remodeling and turnover, and it could be identified as a pathomechanism in bone metabolism diseases.5 Based on bioinformatics analysis of the microarray data set, this study found that bone metabolism altered in hypothyroidism status. In this study, we marked ADIPOQ, SIRT1, PPAR-ɑ, DNMT1, NCOR1, and CTNNB1 as hub genes in the bone metabolism network based on in-silico analysis. Enrichment of hub genes pinpointed adipocytokine, oxidative stress, peroxisome, thyroid hormones, PPAR pathway, Wnt/B-catenin, primary immune deficiency, and metabolism as the significant molecular signaling pathways related to hub genes in bone metabolism. Text mining about pivotal molecular signaling pathways established new strategies to maintain homeostasis of bone metabolism. Stunes et al illustrated that activation of PPAR-ɑ is associated with increased bone mass and positively affects skeletal function.59 SIRT1 or Sirtuin 1, one member of the Sirtuins family, is identified as the most conserved deacetylase in mammals with transcription factor action in several physiological processes such as oxidative and antioxidative mechanism, metabolism, mitochondrial biogenesis, autophagy, apoptosis, and pathological status.60 Growing evidence has proved that SIRT1 with anti-inflammatory action could suppress inflammation in multiple tissues.60,61
On the other hand, NF-κB is specified as the master transcriptional factor in cellular biology and physiology. The NF-κB agent could control DNA transcription, cytokines production, and cell survival. NF-κB is activated due to oxidative stress, free radicals, cytokine production, lipoprotein peroxidation, and led to inflammatory factors production.62 Due to that translocation and activation of the NF-κB agent relies on acetylation, SIRT1 activity prevented NF-κB transcriptional activity suppressing inflammation. Moreover, SIRT1 drives cell proliferation and regulates cell differentiation by deacetylates of β-catenin.63 inhibition of the Wnt signaling pathway leads to protein aggregation, generation of early cells, and accumulation of abnormal extracellular bodies.5 In addition, ADIPOQ is closely associated with bone metabolism, and evidence indicates that ADIPOQ mediates bone metabolism through three pathways: autocrine, paracrine, and insulin signaling pathways.64
We found that the concentration of DDk1 and sclerostin, identified as endogenous inhibitors of Wnt/beta-catenin, increased in hypothyroidism. These data agreed with Tsourdi and colleagues, which revealed that the concentration of the DDk1 and sclerostin increased, and the Wnt/beta-catenin signaling pathway was dysregulated in hypothyroidism conditions.5 Moreover, we demonstrated that the relative expression of Adipoq, Dnmt1, Sirt-1, Ppar-ɑ, Ncor1, and Ctnnb1 genes was disturbed in hypothyroid mice. Increasing Wnt/beta-catenin endogenous inhibitors concentration in hypothyroidism conditions is a retrieval mechanism for modulating bone homeostasis. Thyroid hormones (THs) are anabolic actors in skeletal cell development, bone mass retention, effective healing of bone fracture, and bone metabolism process.65 Based on growing evidence, THs could manage bone turnover, osteoblastogenesis, and osteoclastogenesis. Moreover, PTU is prescribed as a chemical medication for suppressing the hyperactivity of thyroid hormones. In our study, PTU was utilized for hypothyroidism phenotype induction. Hence, the current study suggested that aerobic physical activity and polyphenolic cocktails might suppress PTU function and inhibit the hypothyroidism phenotype, thus offsetting bone metabolism in the hypothyroidism status. T3 induced markers of hypertrophic chondrocyte differentiation, including alkaline phosphatase and collagen X expression in primary growth-plate chondrocyte cultures and enhanced cartilage matrix mineralization. Data mining about chlorogenic acid compound effects has shown that this phytochemical formula is pivotal as the potential antioxidant in scavenging free radicals. Recent studies on the anti-inflammatory effects of honey's common phenolic compounds have found that chlorogenic acid, one of the significant phenolic chemicals in honey, suppresses proinflammatory cytokines tumor necrosis factor (TNFα) and interleukin (IL)-1 expression in mice with neurotoxic intoxicated by MPTP while increasing IL-10 expression.66
Additionally, Coumaric acid has been shown to inhibit IL-8 production in cigarette smoke extract-stimulated A549 cells.67 The effectiveness of Caffeic Acid was shown by Jung et al. The synthesis of iNOS and COX-2 was inhibited, while the production of haem oxygenase-1 was increased in response to LPS.68 Polyphenol compounds mentioned in previous studies with anti-inflammatory and antioxidative stress properties are present in the polyphenol cocktail used in our study.
According to the experimental analysis, we indicated that administering a phytochemicals cocktail and exercise training could modify the T3, T4, and TSH concentrations, which led to modulate bone remodeling. Our data indicated that phytochemicals cocktail and exercise training regulated the relative expression of bone remodeling markers, including Alp, Osx, Sost, Trap, and other biomarkers such as DDk1 and sclerostin. The research by L.G. Rao et focused on examining a combination of flavonoids, including quercetin, apigenin, kaempferol, and luteolin. Based on the data, consuming these supplementation reduced the risk of osteoporosis in postmenopausal women.69 It has been shown in the study by Rao et al. that the observed enhancement is concomitant with reductions in the levels of reactive oxygen species, namely hydrogen peroxide (H2O2).70
In summary, the findings of Rao et al research indicated that the addition of antioxidants such as lycopene and polyphenols may serve as a preventive measure against the development of osteoporosis, which is attributed to oxidative stress induced by reactive oxygen species (ROS). The findings from in vitro investigations conducted on osteoblasts and osteoclasts, as well as animal intervention studies, epidemiological studies, and clinical intervention studies, provide substantiation for the potential utilization of lycopene and polyphenols as supplementary or adjunctive therapeutic agents alongside established pharmaceuticals-approved for the prevention or treatment of osteoporosis in postmenopausal women.70 Polyphenols are well-recognized for mitigating chronic ailments, including malignancies, diabetes, cardiovascular illnesses, neurological disorders, and osteoporosis. Horcajada and Offord reviewed the anabolic function of phytonutrients, with a particular focus on polyphenols and bone health.71 Trzeciakiewicz et al examined how polyphenols affect osteoblast function and their interaction with osteoclasts.72
Vani and co-workers demonstrated that consuming a phytochemical cocktail might ameliorate the antioxidant status and decrease the ischaemic brain.17 These results showed that the phytochemicals cocktail increased the catalase activities, superoxide dismutase, and glutathione levels. In addition, Mostafavi et alindicated that 0.4 mg/kg phytochemicals cocktail had a protective effect on liver function compared to 0.8 mL/kg, 1.6 mL/kg, and 3.2 mL/kg in the mice model.73
Due to the role of oxidative stress and inflammation in the development of osteoporosis, it is plausible to suggest that a functional diet capable of mitigating these mechanisms might serve as a viable preventive intervention to prevent the deterioration of bone quality.74 In a study by Alghadir et al, the objective was to evaluate the impact of a 12-week standardized aerobic training program on the osteogenic indexes and bone markers among individuals without any underlying health conditions. The study specifically examined the levels of blood bone formation indicators, including osteocalcin, alkaline phosphatase, deoxypyridinoline (a marker for bone resorption), and serum calcium. The findings indicated that the 12-week period of moderate AE resulted in a notable enhancement in several bone metabolism indicators, such as Serum bone-specific alkaline phosphatase, serum osteocalcin, serum-free Calcium, and bone mineral density, across all participants. The data mentioned above indicate that engaging in aerobic activity over an extended period is linked to a reduction in bone resorption and maintenance rate or an increase in bone production.75
On the other hand, Zhang et aldemonstrated that two weeks of wheel-running exercise training modulated the trabecular bone, cortical bone, growth plate, and articular cartilage.76 Moreover, wheel-running exercise training modified the osteogenic biomarker and increased the relative expression of Pgc-1 as a co-activator proliferator-activated receptor, which could have a key role in regulating metabolism. Interestingly, Kim and colleagues indicated that physical activity might increase bone resorption, but after that, the osteoclast biomarker was reduced, and bone remodeling was balanced.77 The study conducted by Ooi et al examined the synergistic impact of an eight-week jumping exercise regimen and honey supplementation on bone health in a rat model.78 The rats were orally administered honey, without specification of its kind and content, at a dose of 1 g/kg body weight/day by oral gavage 30 min before the jumping activity. Every rat was exposed to the workout regimen for five minutes daily, five days per week. The combined regimen had better benefits on the tibial bone geometry and mechanical characteristics in rats than the jumping exercise or honey supplementation alone.78
Another research conducted by Tavafzadeh et al had a length of either 8 or 16 weeks.79 The findings of this study revealed significant enhancements in many physiological parameters among young female rats, including tibial wet and fat-free dry weights, tibial and femoral maximal load, tibial midshaft minimum diameter, and femoral midshaft maximum diameter.79
Moreover, we revealed that the expression level of the Adipoq, Dnmt1, Sirt-1, Ppar-ɑ, Ncor1, and Ctnnb1 genes was modified by phytochemicals cocktail and exercise training. Based on the artificial intelligence analysis, we found that oxidative stress, peroxisomes signaling pathway, thyroid hormones, PPARs pathway, Wnt/B-catenin, and metabolism are crucial in the pathogenesis of bone in hypothyroidism status. Moreover, we have shown that administration of physical activity and 0.4 mg/kg phytochemicals cocktail could improve the distribution of osteoblasts and osteoclasts and offset bone metabolism in hypothyroidism in mice via oxidative stress, peroxisomes signaling pathway, thyroid hormones, PPARs pathway, Wnt/B-catenin, and metabolism.
The connection between the phytochemical cocktail and AE lies in their complementary effects on metabolic pathways and cellular mechanisms that influence bone turnover.80 The phytochemical cocktail contains bioactive compounds known to exert antioxidant and anti-inflammatory properties, which can mitigate oxidative stress—an important factor in bone health.81 AE, on the other hand, is known to stimulate mechanical loading on bones, promoting osteogenesis through various signaling pathways, including the Wnt/β-catenin and RANK/RANKL pathways.82 Hence, emerging study have indicated the combined phytochemical cocktail and AE have synergistic mechanisms.43,61,80 When combined, these approaches may amplify the activation of these pathways, leading to a more robust enhancement of bone turnover. Moreover, both interventions may modulate endocrine factors, such as thyroid hormones, which are crucial in bone metabolism, especially in hypothyroid conditions.83 The phytochemical cocktail may enhance thyroid hormone sensitivity or levels, while AE can improve overall hormonal balance and metabolic health, further supporting bone remodeling processes.22 In addition, exercise has been shown to produce beneficial adaptations in inflammatory profiles.43 When combined with the anti-inflammatory effects of the phytochemical cocktail, the result may be a more favorable environment for bone health, reducing the negative effects of chronic inflammation observed in hypothyroid states.84 Ultimately, understanding how these two interventions interact helps clarify their joint contributions toward restoring bone integrity and function in hypothyroid mice, which could have broader implications for therapeutic strategies in bone health management.
Epigenetic modification might play a crucial role in the pathogenesis of disorders and diseases and might manifest as potential biomarkers in prognosis and diagnosis.85 Based on in silico prediction, the present study highlighted lncRNAs as the pivotal regulators in gene expression. We predicted long non-coding RNAs HOTAIR, MEG3, H19, and TUG1 based on the target gene algorithm and found that TUG1 could regulate the CTNNB1 and SIRT1 gene expression. On the other hand, we found that MEG3 and H19 could be targeted and regulated by DNMT1. Also, we recognized HOTAIR as regulatory factors with roles in histone modification, sponge, and interaction with the proteins in the Wnt/β-catenin signaling pathway.
Yu and colleagues reported that TUG1 scavenges miR-204-5p to enhance osteoblast development in aortic valve calcification by upregulating Runx2.86 We suggested that the upregulation of TUG1 leads to cell differentiation via targeting RUNX2. Furthermore, TUG1, with a positive effect on CTNNB1, could regulate cancer progression via interacting with Wnt/beta-catenin signaling.87 On the other hand, Xiang et al have shown that HOTAIR targeted DNMT1 and has a positive effect on cell growth and a negative effect on the apoptosis process via interaction mechanism with protein.88 Meanwhile, upregulation of HOTAIR negatively affects CTNNB1 and enhances apoptosis by inactivating beta-catenin in lung cancer cells.89 Also, we recognized HOTAIR as regulatory factors with roles in histone modification, sponge, and interaction with the proteins in the Wnt/β-catenin signaling pathway. Moreover, MEG3 regulates cell proliferation, and the Wnt/β-catenin signaling pathway by targeting DNMT1 and CTNNB1.90 The long non-coding RNA H19 encourages cell proliferation in colorectal cancer by competitively binding to miR-200a and inhibiting beta-Catenin expression.91 Notably, Li et al indicated that overexpression of DNMT1 triggers hypermethylation H19 in Disuse osteoporosis rats. Overexpression of H19 is associated with osteogenic differentiation and bone formation.77
Here, we showed that relative expression of HOTAIR, TUG1, and H19 are increased in hypothyroidism mice, and MEG3 expression declined in hypothyroidism status. Thus, we recommended that administering physical activity and a 0.4 mg/kg Phytochemicals cocktail led to an offsetting mechanism in bone metabolism disruptions induced by hypothyroidism. Although bioinformatics and molecular docking provide substantial insights, the lack of molecular dynamics simulations constrains the precision of predictions about ligand-protein interactions. Future analysis requires dynamic simulations to validate these conclusions. Future studies should consider these limitations and explore adjustments in simulation parameters and control selections to enhance the reliability and interpretability of the results.
Conclusion
The interaction of AE and a 0.4 mg/kg Phytochemicals cocktail could modulate Metabolism, Oxidative stress, and Wnt signaling pathway in the bone of hypothyroid mice.
Research Highlights
What is the current knowledge?
What is new here?
-
DA and AE could have a synergistic effect on the rehabilitation of HYPO.
-
DA and AE modulated the vital hub genes network in bone tissue.
-
Molecular diagnosis of hypothyroidism effect on bone tissue via biomarkers.
Competing Interests
There is no competing of interest to disclose
Ethical Approval
This study was performed and approved based on all the protocols of laboratory animals by the Ethics Committee of Islamic Azad University, Khorasgan Branch (IR.IAU.KHUISF.REC.1400.231).
Acknowledgements
We thank our colleagues for their association and helpful discussions in this study.
References
- Acharya B, Chaijaroenkul W, Na-Bangchang K. Therapeutic potential and pharmacological activities of β-eudesmol. Chem Biol Drug Des 2021; 97:984-96. doi: 10.1111/cbdd.13823 [Crossref] [ Google Scholar]
- Robson H, Siebler T, Stevens DA, Shalet SM, Williams GR. Thyroid hormone acts directly on growth plate chondrocytes to promote hypertrophic differentiation and inhibit clonal expansion and cell proliferation. Endocrinology 2000; 141:3887-97. doi: 10.1210/endo.141.10.7733 [Crossref] [ Google Scholar]
- Varga F, Rumpler M, Zoehrer R, Turecek C, Spitzer S, Thaler R. T3 affects expression of collagen I and collagen cross-linking in bone cell cultures. Biochem Biophys Res Commun 2010; 402:180-5. doi: 10.1016/j.bbrc.2010.08.022 [Crossref] [ Google Scholar]
- Govindan R, El-Sherbiny M, Ibraheem KM, Narasimhan S, Salama ME, Ahmad F. Thyroid-stimulating hormone favors Runx2-mediated matrix mineralization in HOS and SaOS2 cells: an in vitro and in silico approach. Molecules 2022; 27:613. doi: 10.3390/molecules27030613 [Crossref] [ Google Scholar]
- Tsourdi E, Rijntjes E, Köhrle J, Hofbauer LC, Rauner M. Hyperthyroidism and hypothyroidism in male mice and their effects on bone mass, bone turnover, and the Wnt inhibitors sclerostin and Dickkopf-1. Endocrinology 2015; 156:3517-27. doi: 10.1210/en.2015-1073 [Crossref] [ Google Scholar]
- Pai SG, Carneiro BA, Mota JM, Costa R, Leite CA, Barroso-Sousa R. Wnt/beta-catenin pathway: modulating anticancer immune response. J Hematol Oncol 2017; 10:101. doi: 10.1186/s13045-017-0471-6 [Crossref] [ Google Scholar]
- Mosekilde L, Eriksen EF, Charles P. Effects of thyroid hormones on bone and mineral metabolism. Endocrinol Metab Clin North Am 1990; 19:35-63. doi: 10.1016/S0889-8529(18)30338-4 [Crossref] [ Google Scholar]
- Hajibabaie F, Abedpoor N, Mohamadynejad P. Types of cell death from a molecular perspective. Biology (Basel) 2023; 12:1426. doi: 10.3390/biology12111426 [Crossref] [ Google Scholar]
- Rivkees SA, Bode HH, Crawford JD. Long-term growth in juvenile acquired hypothyroidism: the failure to achieve normal adult stature. N Engl J Med 1988; 318:599-602. doi: 10.1056/nejm198803103181003 [Crossref] [ Google Scholar]
- Dallakyan S, Olson AJ. Small-molecule library screening by docking with PyRx. Methods Mol Biol 2015; 1263:243-50. doi: 10.1007/978-1-4939-2269-7_19 [Crossref] [ Google Scholar]
- O'Shea PJ, Kim DW, Logan JG, Davis S, Walker RL, Meltzer PS. Advanced bone formation in mice with a dominant-negative mutation in the thyroid hormone receptor β gene due to activation of Wnt/β-catenin protein signaling. J Biol Chem 2012; 287:17812-22. doi: 10.1074/jbc.M111.311464 [Crossref] [ Google Scholar]
- Hajibabaie F, Abedpoor N, Assareh N, Tabatabaiefar MA, Shariati L, Zarrabi A. The importance of SNPs at miRNA binding sites as biomarkers of gastric and colorectal cancers: a systematic review. J Pers Med 2022; 12:456. doi: 10.3390/jpm12030456 [Crossref] [ Google Scholar]
- Zhang X, Wang W, Zhu W, Dong J, Cheng Y, Yin Z. Mechanisms and functions of long non-coding RNAs at multiple regulatory levels. Int J Mol Sci 2019; 20:5573. doi: 10.3390/ijms20225573 [Crossref] [ Google Scholar]
- Hajibabaie F, Abedpoor N, Haghjooy Javanmard S, Hasan A, Sharifi M, Rahimmanesh I. The molecular perspective on the melanoma and genome engineering of T-cells in targeting therapy. Environ Res 2023; 237:116980. doi: 10.1016/j.envres.2023.116980 [Crossref] [ Google Scholar]
- Mainardi T, Kapoor S, Bielory L. Complementary and alternative medicine: herbs, phytochemicals and vitamins and their immunologic effects. J Allergy Clin Immunol 2009; 123:283-94. doi: 10.1016/j.jaci.2008.12.023 [Crossref] [ Google Scholar]
- Hossiniyan SA, Ghalamfarsa F, Rostami-Yasuj S, Salehpoor Z, Latifpour M, Hasanzadeh S. Chemical composition and cytotoxic activity of the essential oil from the aerial parts of Dorema aucheri. J Herbmed Pharmacol 2021; 10:344-50. doi: 10.34172/jhp.2021.40 [Crossref] [ Google Scholar]
- Rasouli Vani J, Mohammadi MT, Sarami Foroshani M, Rezazade E. Evaluation of the neuroprotective and antioxidant effects of Dorema aucheri extract on cerebral ischaemia-reperfusion injury in rats. Pharm Biol 2019; 57:255-62. doi: 10.1080/13880209.2019.1597132 [Crossref] [ Google Scholar]
- Kaviani E, Hajibabaie F, Abedpoor N, Safavi K, Ahmadi Z, Karimy A. System biology analysis to develop diagnostic biomarkers, monitoring pathological indexes, and novel therapeutic approaches for immune targeting based on maggot bioactive compounds and polyphenolic cocktails in mice with gastric cancer. Environ Res 2023; 238:117168. doi: 10.1016/j.envres.2023.117168 [Crossref] [ Google Scholar]
- Hajibabaie F, Abedpoor N, Safavi K, Taghian F. Natural remedies medicine derived from flaxseed (secoisolariciresinol diglucoside, lignans, and α-linolenic acid) improve network targeting efficiency of diabetic heart conditions based on computational chemistry techniques and pharmacophore modeling. J Food Biochem 2022; 46:e14480. doi: 10.1111/jfbc.14480 [Crossref] [ Google Scholar]
- Hajibabaie F, Abedpoor N, Taghian F, Safavi K. A cocktail of polyherbal bioactive compounds and regular mobility training as senolytic approaches in age-dependent Alzheimer's: the in-silico analysis, lifestyle intervention in old age. J Mol Neurosci 2023; 73:171-84. doi: 10.1007/s12031-022-02086-8 [Crossref] [ Google Scholar]
- Abedpoor N, Taghian F, Hajibabaie F. Cross brain-gut analysis highlighted hub genes and lncRNA networks differentially modified during leucine consumption and endurance exercise in mice with depression-like behaviors. Mol Neurobiol 2022; 59:4106-23. doi: 10.1007/s12035-022-02835-1 [Crossref] [ Google Scholar]
- Abedpoor N, Taghian F, Hajibabaie F. Physical activity ameliorates the function of organs via adipose tissue in metabolic diseases. Acta Histochem 2022; 124:151844. doi: 10.1016/j.acthis.2022.151844 [Crossref] [ Google Scholar]
- Kim K. Effects of exercise training on thyroid function, body composition, physical fitness, and blood lipid profiles in subclinical hypothyroidism subjects. Korean J Phys Educ 2012; 51:385-94. [ Google Scholar]
- Singh S, Panda V, S S, Dande P. Protective effect of a polyherbal bioactive fraction in propylthiouracil-induced thyroid toxicity in ratsby modulation of the hypothalamic-pituitary-thyroid and hypothalamic-pituitary-adrenal axes. Toxicol Rep 2020; 7:730-42. doi: 10.1016/j.toxrep.2020.06.002 [Crossref] [ Google Scholar]
- Gao Y, Wang J, Zhou Y, Sheng S, Qian SY, Huo X. Evaluation of serum CEA, CA19-9, CA72-4, CA125 and ferritin as diagnostic markers and factors of clinical parameters for colorectal cancer. Sci Rep 2018; 8:2732. doi: 10.1038/s41598-018-21048-y [Crossref] [ Google Scholar]
- Rahbar Saadat Y, Pourseif MM, Zununi Vahed S, Barzegari A, Omidi Y, Barar J. Modulatory role of vaginal-isolated Lactococcus lactis on the expression of miR-21, miR-200b, and TLR-4 in CAOV-4 cells and in silico revalidation. Probiotics Antimicrob Proteins 2020; 12:1083-96. doi: 10.1007/s12602-019-09596-9 [Crossref] [ Google Scholar]
- Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res 2003; 13:2498-504. doi: 10.1101/gr.1239303 [Crossref] [ Google Scholar]
- Johnson E. Gephi-Network Analysis and Visualization. Miami University; 2019. Available from: http://hdl.handle.net/2374.MIA/6337.
- Piñero J, Ramírez-Anguita JM, Saüch-Pitarch J, Ronzano F, Centeno E, Sanz F. The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Res 2020; 48:D845-55. doi: 10.1093/nar/gkz1021 [Crossref] [ Google Scholar]
- Oliveros JC. VENNY. An Interactive Tool for Comparing Lists with Venn's Diagrams. 2007. Available from: https://bioinfogp.cnb.csic.es/tools/venny/index.html.
- Bu D, Luo H, Huo P, Wang Z, Zhang S, He Z. KOBAS-i: intelligent prioritization and exploratory visualization of biological functions for gene enrichment analysis. Nucleic Acids Res 2021; 49:W317-25. doi: 10.1093/nar/gkab447 [Crossref] [ Google Scholar]
- Xie Z, Bailey A, Kuleshov MV, Clarke DJ, Evangelista JE, Jenkins SL. Gene set knowledge discovery with Enrichr. Curr Protoc 2021; 1:e90. doi: 10.1002/cpz1.90 [Crossref] [ Google Scholar]
- Buniello A, MacArthur JA, Cerezo M, Harris LW, Hayhurst J, Malangone C. The NHGRI-EBI GWAS Catalog of published genome-wide association studies, targeted arrays and summary statistics 2019. Nucleic Acids Res 2019; 47:D1005-12. doi: 10.1093/nar/gky1120 [Crossref] [ Google Scholar]
- Zhao H, Shi J, Zhang Y, Xie A, Yu L, Zhang C. LncTarD: a manually-curated database of experimentally-supported functional lncRNA-target regulations in human diseases. Nucleic Acids Res 2020; 48:D118-26. doi: 10.1093/nar/gkz985 [Crossref] [ Google Scholar]
- Bao Z, Yang Z, Huang Z, Zhou Y, Cui Q, Dong D. LncRNADisease 20: an updated database of long non-coding RNA-associated diseases. Nucleic Acids Res 2019; 47:D1034-7. doi: 10.1093/nar/gky905 [Crossref] [ Google Scholar]
- Ning L, Cui T, Zheng B, Wang N, Luo J, Yang B. MNDR v30: mammal ncRNA-disease repository with increased coverage and annotation. Nucleic Acids Res 2021; 49:D160-4. doi: 10.1093/nar/gkaa707 [Crossref] [ Google Scholar]
- Ma L, Cao J, Liu L, Du Q, Li Z, Zou D. LncBook: a curated knowledgebase of human long non-coding RNAs. Nucleic Acids Res 2019; 47:D128-34. doi: 10.1093/nar/gky960 [Crossref] [ Google Scholar]
- Volders PJ, Anckaert J, Verheggen K, Nuytens J, Martens L, Mestdagh P. LNCipedia 5: towards a reference set of human long non-coding RNAs. Nucleic Acids Res 2019; 47:D135-9. doi: 10.1093/nar/gky1031 [Crossref] [ Google Scholar]
- Abedpoor N, Taghian F, Ghaedi K, Niktab I, Safaeinejad Z, Rabiee F. PPARγ/Pgc-1α-Fndc5 pathway up-regulation in gastrocnemius and heart muscle of exercised, branched chain amino acid diet fed mice. Nutr Metab (Lond) 2018; 15:59. doi: 10.1186/s12986-018-0298-3 [Crossref] [ Google Scholar]
- Fukunaga T, Iwakiri J, Ono Y, Hamada M. LncRRIsearch: a web server for lncRNA-RNA interaction prediction integrated with tissue-specific expression and subcellular localization data. Front Genet 2019; 10:462. doi: 10.3389/fgene.2019.00462 [Crossref] [ Google Scholar]
- Chang L, Zhou G, Soufan O, Xia J. miRNet 20: network-based visual analytics for miRNA functional analysis and systems biology. Nucleic Acids Res 2020; 48:W244-51. doi: 10.1093/nar/gkaa467 [Crossref] [ Google Scholar]
- Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM, Meng EC. UCSF Chimera--a visualization system for exploratory research and analysis. J Comput Chem 2004; 25:1605-12. doi: 10.1002/jcc.20084 [Crossref] [ Google Scholar]
- Abedpoor N, Taghian F, Jalali Dehkordi K, Safavi K. Sparassis latifolia and exercise training as complementary medicine mitigated the 5-fluorouracil potent side effects in mice with colorectal cancer: bioinformatics approaches, novel monitoring pathological metrics, screening signatures, and innovative management tactic. Cancer Cell Int 2024; 24:141. doi: 10.1186/s12935-024-03328-y [Crossref] [ Google Scholar]
- Zhou Y, Gao Y, Xu C, Shen H, Tian Q, Deng HW. A novel approach for correction of crosstalk effects in pathway analysis and its application in osteoporosis research. Sci Rep 2018; 8:668. doi: 10.1038/s41598-018-19196-2 [Crossref] [ Google Scholar]
- Tavares SJ, Lima V. Bone anti-resorptive effects of coumarins on RANKL downstream cellular signaling: a systematic review of the literature. Fitoterapia 2021; 150:104842. doi: 10.1016/j.fitote.2021.104842 [Crossref] [ Google Scholar]
- Zhang P, Ye J, Dai J, Wang Y, Chen G, Hu J. Gallic acid inhibits osteoclastogenesis and prevents ovariectomy-induced bone loss. Front Endocrinol (Lausanne) 2022; 13:963237. doi: 10.3389/fendo.2022.963237 [Crossref] [ Google Scholar]
- Afzal A, Oriqat G, Akram Khan M, Jose J, Afzal M. Chemistry and biochemistry of terpenoids from Curcuma and related species. J Biol Act Prod Nat 2013; 3:1-55. doi: 10.1080/22311866.2013.782757 [Crossref] [ Google Scholar]
- Yu S, Long Y, Li D, Shi A, Deng J, Ma Y. Natural essential oils efficacious in internal organs fibrosis treatment: mechanisms of action and application perspectives. Pharmacol Res 2022; 182:106339. doi: 10.1016/j.phrs.2022.106339 [Crossref] [ Google Scholar]
- Mao W, Huang G, Chen H, Xu L, Qin S, Li A. Research progress of the role of anthocyanins on bone regeneration. Front Pharmacol 2021; 12:773660. doi: 10.3389/fphar.2021.773660 [Crossref] [ Google Scholar]
- Nguyen LV, Nguyen KD, Ma CT, Nguyen QT, Nguyen HT, Yang DJ. p-Coumaric acid enhances hypothalamic leptin signaling and glucose homeostasis in mice via differential effects on AMPK activation. Int J Mol Sci 2021; 22:1431. doi: 10.3390/ijms22031431 [Crossref] [ Google Scholar]
- Lee JH, Chung YH, Kim HH, Bang JS, Jung TW, Park T. p-Coumaric acid stimulates longitudinal bone growth through increasing the serum production and expression levels of insulin-like growth factor 1 in rats. Biochem Biophys Res Commun 2018; 505:1103-6. doi: 10.1016/j.bbrc.2018.10.046 [Crossref] [ Google Scholar]
- Tolba MF, El-Serafi AT, Omar HA. Caffeic acid phenethyl ester protects against glucocorticoid-induced osteoporosis in vivo: impact on oxidative stress and RANKL/OPG signals. Toxicol Appl Pharmacol 2017; 324:26-35. doi: 10.1016/j.taap.2017.03.021 [Crossref] [ Google Scholar]
- Zhou RP, Lin SJ, Wan WB, Zuo HL, Yao FF, Ruan HB. Chlorogenic acid prevents osteoporosis by Shp2/PI3K/Akt pathway in ovariectomized rats. PLoS One 2016; 11:e0166751. doi: 10.1371/journal.pone.0166751 [Crossref] [ Google Scholar]
- Kwak SC, Lee C, Kim JY, Oh HM, So HS, Lee MS. Chlorogenic acid inhibits osteoclast differentiation and bone resorption by down-regulation of receptor activator of nuclear factor kappa-B ligand-induced nuclear factor of activated T cells c1 expression. Biol Pharm Bull 2013; 36:1779-86. doi: 10.1248/bpb.b13-00430 [Crossref] [ Google Scholar]
- Han D, Gu X, Gao J, Wang Z, Liu G, Barkema HW. Chlorogenic acid promotes the Nrf2/HO-1 anti-oxidative pathway by activating p21(Waf1/Cip1) to resist dexamethasone-induced apoptosis in osteoblastic cells. Free Radic Biol Med 2019; 137:1-12. doi: 10.1016/j.freeradbiomed.2019.04.014 [Crossref] [ Google Scholar]
- Carvalho AM, Heimfarth L, Pereira EW, Oliveira FS, Menezes IR, Coutinho HD. Phytol, a chlorophyll component, produces antihyperalgesic, anti-inflammatory, and antiarthritic effects: possible NFκB pathway involvement and reduced levels of the proinflammatory cytokines TNF-α and IL-6. J Nat Prod 2020; 83:1107-17. doi: 10.1021/acs.jnatprod.9b01116 [Crossref] [ Google Scholar]
- Kim EN, Trang NM, Kang H, Kim KH, Jeong GS. Phytol suppresses osteoclast differentiation and oxidative stress through Nrf2/HO-1 regulation in RANKL-induced RAW2647 cells. Cells 2022; 11:3596. doi: 10.3390/cells11223596 [Crossref] [ Google Scholar]
- Yamaguchi M, Levy RM. β-Caryophyllene promotes osteoblastic mineralization, and suppresses osteoclastogenesis and adipogenesis in mouse bone marrow cultures in vitro. Exp Ther Med 2016; 12:3602-6. doi: 10.3892/etm.2016.3818 [Crossref] [ Google Scholar]
- Stunes AK, Westbroek I, Gustafsson BI, Fossmark R, Waarsing JH, Eriksen EF. The peroxisome proliferator-activated receptor (PPAR) alpha agonist fenofibrate maintains bone mass, while the PPAR gamma agonist pioglitazone exaggerates bone loss, in ovariectomized rats. BMC Endocr Disord 2011; 11:11. doi: 10.1186/1472-6823-11-11 [Crossref] [ Google Scholar]
- Iyer S, Han L, Bartell SM, Kim HN, Gubrij I, de Cabo R. Sirtuin1 (Sirt1) promotes cortical bone formation by preventing β-catenin sequestration by FoxO transcription factors in osteoblast progenitors. J Biol Chem 2014; 289:24069-78. doi: 10.1074/jbc.M114.561803 [Crossref] [ Google Scholar]
- Rahimi G, Heydari S, Rahimi B, Abedpoor N, Niktab I, Safaeinejad Z. A combination of herbal compound (SPTC) along with exercise or metformin more efficiently alleviated diabetic complications through down-regulation of stress oxidative pathway upon activating Nrf2-Keap1 axis in AGE rich diet-induced type 2 diabetic mice. Nutr Metab (Lond) 2021; 18:14. doi: 10.1186/s12986-021-00543-6 [Crossref] [ Google Scholar]
- Sergi C, Shen F, Liu SM. Insulin/IGF-1R, SIRT1, and FOXOs pathways-an intriguing interaction platform for bone and osteosarcoma. Front Endocrinol (Lausanne) 2019; 10:93. doi: 10.3389/fendo.2019.00093 [Crossref] [ Google Scholar]
- Demory ML, Boerner JL, Davidson R, Faust W, Miyake T, Lee I. Epidermal growth factor receptor translocation to the mitochondria: regulation and effect. J Biol Chem 2009; 284:36592-604. doi: 10.1074/jbc.M109.000760 [Crossref] [ Google Scholar]
- Zhou M, Ning N, Jiang Y, Aschner M, Huang X, Bin X. Correlation of ADIPOQ gene single nucleotide polymorphisms with bone strength index in middle-aged and the elderly of Guangxi Mulam ethnic group. Int J Environ Res Public Health 2021; 18:13034. doi: 10.3390/ijerph182413034 [Crossref] [ Google Scholar]
- Williams GR. Thyroid hormone actions in cartilage and bone. Eur Thyroid J 2013; 2:3-13. doi: 10.1159/000345548 [Crossref] [ Google Scholar]
- Singh SS, Rai SN, Birla H, Zahra W, Kumar G, Gedda MR. Effect of chlorogenic acid supplementation in MPTP-intoxicated mouse. Front Pharmacol 2018; 9:757. doi: 10.3389/fphar.2018.00757 [Crossref] [ Google Scholar]
- da Silva EC, Dos Santos FM, Ribeiro AR, de Souza ST, Barreto E, da Silva Fonseca EJ. Drug-induced anti-inflammatory response in A549 cells, as detected by Raman spectroscopy: a comparative analysis of the actions of dexamethasone and p-coumaric acid. Analyst 2019; 144:1622-31. doi: 10.1039/c8an01887a [Crossref] [ Google Scholar]
- Jung WK, Lee DY, Kim JH, Choi I, Park SG, Seo SK. Anti-inflammatory activity of caffeic acid phenethyl ester (CAPE) extracted from Rhodiola sacra against lipopolysaccharide-induced inflammatory responses in mice. Process Biochem 2008; 43:783-7. doi: 10.1016/j.procbio.2008.03.004 [Crossref] [ Google Scholar]
- Rao V, Balachandran B, Shen H, Logan A, Rao L. In vitro and in vivo antioxidant properties of the plant-based supplement greens + TM. Int J Mol Sci 2011; 12:4896-908. doi: 10.3390/ijms12084896 [Crossref] [ Google Scholar]
- Rao LG, Balachandran B, Rao AV. Polyphenol extract of Greens + TM nutritional supplement stimulates bone formation in cultures of human osteoblast-like SaOS-2 cells. J Diet Suppl 2008; 5:264-82. doi: 10.1080/19390210802414295 [Crossref] [ Google Scholar]
- Horcajada MN, Offord E. Naturally plant-derived compounds: role in bone anabolism. Curr Mol Pharmacol 2012; 5:205-18. doi: 10.2174/1874467211205020205 [Crossref] [ Google Scholar]
- Trzeciakiewicz A, Habauzit V, Horcajada MN. When nutrition interacts with osteoblast function: molecular mechanisms of polyphenols. Nutr Res Rev 2009; 22:68-81. doi: 10.1017/s095442240926402x [Crossref] [ Google Scholar]
- Mostafavi SH, Fazilati M, Mostafavi SA, Vahhabi MR, Mostafavi F, Omidvarinia S. Hepatotoxicity of Dorema aucheri (Bilhar) in albino mice. Arch Iran Med 2013; 16:530-2. [ Google Scholar]
- Kamaruzzaman MA, Chin KY, Mohd Ramli ES. A review of potential beneficial effects of honey on bone health. Evid Based Complement Alternat Med 2019; 2019:8543618. doi: 10.1155/2019/8543618 [Crossref] [ Google Scholar]
- Alghadir AH, Aly FA, Gabr SA. Effect of moderate aerobic training on bone metabolism indices among adult humans. Pak J Med Sci 2014; 30:840-4. doi: 10.12669/pjms.304.4624 [Crossref] [ Google Scholar]
- Zhang J, Valverde P, Zhu X, Murray D, Wu Y, Yu L. Exercise-induced irisin in bone and systemic irisin administration reveal new regulatory mechanisms of bone metabolism. Bone Res 2017; 5:16056. doi: 10.1038/boneres.2016.56 [Crossref] [ Google Scholar]
- Li B, Zhao J, Ma JX, Li GM, Zhang Y, Xing GS. Overexpression of DNMT1 leads to hypermethylation of H19 promoter and inhibition of Erk signaling pathway in disuse osteoporosis. Bone 2018; 111:82-91. doi: 10.1016/j.bone.2018.03.017 [Crossref] [ Google Scholar]
- Ooi FK, Tavafzadeh SS, Hung L, Hung W, He Y. Tibial bone mineral density, geometry, and mechanical properties in response to high-impact exercise and honey supplementation in rats. Asian J Exerc Sport Sci 2014; 11:12-24. [ Google Scholar]
- Tavafzadeh SS, Ooi FK, Chen CK, Sulaiman SA, Hung LK. Bone mechanical properties and mineral density in response to cessation of jumping exercise and honey supplementation in young female rats. Biomed Res Int 2015; 2015:938782. doi: 10.1155/2015/938782 [Crossref] [ Google Scholar]
- Thomas R, Williams M, Aldous J, Wyld K. Multiple biological mechanisms for the potential influence of phytochemicals on physical activity performance: a narrative review. Nutraceuticals 2023; 3:353-65. doi: 10.3390/nutraceuticals3030027 [Crossref] [ Google Scholar]
- Su Z, Yao B, Liu G, Fang J. Polyphenols as potential preventers of osteoporosis: a comprehensive review on antioxidant and anti-inflammatory effects, molecular mechanisms, and signal pathways in bone metabolism. J Nutr Biochem 2024. 123: 109488. doi: 10.1016/j.jnutbio.2023.109488.
- Ma Q, Miri Z, Haugen HJ, Moghanian A, Loca D. Significance of mechanical loading in bone fracture healing, bone regeneration, and vascularization. J Tissue Eng 2023; 14:20417314231172573. doi: 10.1177/20417314231172573 [Crossref] [ Google Scholar]
- Cini G, Carpi A, Mechanick J, Cini L, Camici M, Galetta F. Thyroid hormones and the cardiovascular system: pathophysiology and interventions. Biomed Pharmacother 2009; 63:742-53. doi: 10.1016/j.biopha.2009.08.003 [Crossref] [ Google Scholar]
- Saleh HA, Yousef MH, Abdelnaser A. The anti-inflammatory properties of phytochemicals and their effects on epigenetic mechanisms involved in TLR4/NF-κB-mediated inflammation. Front Immunol 2021; 12:606069. doi: 10.3389/fimmu.2021.606069 [Crossref] [ Google Scholar]
- Hajibabaie F, Kouhpayeh S, Mirian M, Rahimmanesh I, Boshtam M, Sadeghian L. MicroRNAs as the actors in the atherosclerosis scenario. J Physiol Biochem 2020; 76:1-12. doi: 10.1007/s13105-019-00710-7 [Crossref] [ Google Scholar]
- Yu C, Li L, Xie F, Guo S, Liu F, Dong N. LncRNA TUG1 sponges miR-204-5p to promote osteoblast differentiation through upregulating Runx2 in aortic valve calcification. Cardiovasc Res 2018; 114:168-79. doi: 10.1093/cvr/cvx180 [Crossref] [ Google Scholar]
- Liang S, Zhang S, Wang P, Yang C, Shang C, Yang J. LncRNA, TUG1 regulates the oral squamous cell carcinoma progression possibly via interacting with Wnt/β-catenin signaling. Gene 2017; 608:49-57. doi: 10.1016/j.gene.2017.01.024 [Crossref] [ Google Scholar]
- Xiang S, Zou P, Tang Q, Zheng F, Wu J, Chen Z. HOTAIR-mediated reciprocal regulation of EZH2 and DNMT1 contribute to polyphyllin I-inhibited growth of castration-resistant prostate cancer cells in vitro and in vivo. Biochim Biophys Acta Gen Subj 2018; 1862:589-99. doi: 10.1016/j.bbagen.2017.12.001 [Crossref] [ Google Scholar]
- Chen J, Shen Z, Zheng Y, Wang S, Mao W. Radiotherapy induced Lewis lung cancer cell apoptosis via inactivating β-catenin mediated by upregulated HOTAIR. Int J Clin Exp Pathol 2015; 8:7878-86. [ Google Scholar]
- Xia Y, He Z, Liu B, Wang P, Chen Y. Downregulation of Meg3 enhances cisplatin resistance of lung cancer cells through activation of the WNT/β-catenin signaling pathway. Mol Med Rep 2015; 12:4530-7. doi: 10.3892/mmr.2015.3897 [Crossref] [ Google Scholar]
- Yang W, Ning N, Jin X. The lncRNA H19 promotes cell proliferation by competitively binding to miR-200a and derepressing β-catenin expression in colorectal cancer. Biomed Res Int 2017; 2017:2767484. doi: 10.1155/2017/2767484 [Crossref] [ Google Scholar]