Bioimpacts. 16:31018.
doi: 10.34172/bi.31018
Original Article
Low-level lasers can improve spermatogenesis in infertile mice by targeting molecular pathways
Habib Tajalli Conceptualization, Funding acquisition, Methodology, Supervision, Validation, Writing – review & editing, 1, 2, * 
Masoud Maleki Conceptualization, Methodology, Supervision, Validation, Writing – review & editing, 3, 4
Fatemeh Firouzi Amoudizaj Data curation, Investigation, Methodology, Project administration, Resources, Writing – original draft, Writing – review & editing, 3, 4
Zahra Akbarpour Formal analysis, Investigation, Methodology, Project administration, Writing – original draft, Writing – review & editing, 3, 4
Esmail Safavi Conceptualization, Supervision, Validation, Writing – review & editing, 4, 5
Reza Shahi Formal analysis, Resources, Writing – review & editing, 6
Alireza Sotoudeh Investigation, Methodology, Resources, Writing – review & editing, 1, 2
Ahmad Babazadeh Bedoustani Investigation, Resources, Writing – review & editing, 4, 7
Author information:
1Research Center of Biophotonics, Tabriz Branch, Islamic Azad University, Tabriz, Iran
2Department of Physics, Tabriz Branch, Islamic Azad University, Tabriz, Iran
3Department of Biology, Tabriz Branch, Islamic Azad University, Tabriz, Iran
4Research Center of Biotechnology, Tabriz Branch, Islamic Azad University, Tabriz, Iran
5Department of Basic Sciences, Faculty of Veterinary Medicine, Tabriz Branch, Azad Islamic University, Tabriz, Iran
6Department of Mathematics, Tabriz Branch, Islamic Azad University, Tabriz, Iran
7Department of Biology, Faculty of Natural Science, University of Tabriz, Tabriz, Iran
Abstract
Introduction:
Infertility affects approximately 15% of couples worldwide, with male factors accounting for approximately half of the cases. Low-level laser therapy (LLLT) has been increasingly considered in modern medicine due to its high efficacy, ease of use, and lack of side effects. Evidence suggests that this method can prevent DNA damage in cells and activate key genes related to fertility. This study aimed to investigate the effects of LLLT on sperm production in azoospermic mice using in vitro and in vivo experimental models.
Methods:
Adult male NMRI mice (8–9 weeks old, 30–35 g) were divided into the negative control (healthy), positive control (azoospermia via intraperitoneal busulfan, 30 mg/kg), and experimental (azoospermia with 808 nm LLLT at 8 J/cm2) groups (8 mice per group) for in vivo experiments. For the in vitro part, spermatogonial stem cells were cultured from 6 azoospermic mice under control conditions or treated with laser (808 nm LLLT at 4 J/cm2). Morphological examination and real-time polymerase chain reaction were used to assess testicular structure and expression of several genes, such as deleted in azoospermia-like (DAZL), G protein-coupled receptor 125 (GPR125), synaptonemal complex protein 3 (SYCP3), DEAD-box helicase 4 (VASA/DDX4), protamine (PRM), acrosin (ACR), and tripartite motif containing 36 (Haprin/TRIM36).
Results:
In vivo, LLLT increased VASA expression, improved germ cell activity, and increased sperm production compared with untreated azoospermic control groups, although these changes were not statistically significant. In vitro, 4 J/cm2 radiation modulated several genes related to spermatogenesis, supporting its role in germ cell differentiation.
Conclusion:
LLLT with a wavelength of 808 nm could improve spermatogenesis and sperm production in a mouse model of busulfan-induced azoospermia in vivo and in vitro. These results demonstrated its potential as a supportive treatment for male infertility.
Graphical Abstract
Keywords: Spermatogenesis, Infertility, Azoospermia, Laser therapy
Copyright and License Information
© 2026 The Author(s).
This work is published by BioImpacts as an open access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (
http://creativecommons.org/licenses/by-nc/4.0/). Non-commercial uses of the work are permitted, provided the original work is properly cited.
Funding Statement
This project received financial support from Islamic Azad University of Tabriz.
Introduction
Research indicates that approximately 8-12% of couples of reproductive age globally have infertility issues, with male factors contributing to 50% of these instances.1,2 Azoospermia and oligospermia represent some of the most severe forms of male infertility, impacting 20–30% of patients.3 Azoospermia can be caused by various conditions, such as bacterial infections, reproductive system malignancies, and genetic syndromes (e.g., Prader-Willi syndrome).4 Recent studies have suggested advanced treatment methods, including hormone replacement therapy and intracytoplasmic sperm injection, as potential solutions for male infertility. However, current methods seem to be less effective and have negative effects on azoospermic patients, especially those with nonobstructive azoospermia. Therefore, there is a need for an effective medical treatment plan to minimize side effects.3,5
Recent years have witnessed the consideration of laser therapy, a commonly used method in modern medicine, for the treatment of male infertility due to its high effectiveness, simplicity of use, and lack of adverse effects6,7 Laser therapy is a non-invasive approach that is able to activate various cellular mechanisms and induce new biological responses.8,9 When applied in vivo at the appropriate wavelengths and optimal energy doses, laser therapy can produce a wide range of molecular, cellular, and tissue-level effects.10-14
The effects of laser radiation on sperm function have been extensively studied in animal models. The research results on lasers with different powers, various radiation energies, and different radiation durations revealed that lasers with low power and energy density have the most positive effect on sperm quantitative and qualitative indicators.8-16 Cellular function through low-level laser therapy (LLLT) is mainly caused by stimulating adenosine triphosphate production and reducing oxidative stress. These effects depend on the dose, intensity, and duration of radiation. According to these studies, LLLT can increase human sperm motility.17 Gabel et al., investigating the effects of LLLT on the function, motility, and DNA integrity of human sperm, reported that the effects of laser depend on the radiation dose and duration and can lead to a fourfold increase in adenosine triphosphate production in sperm compared to the control group.17
Previous studies have demonstrated that LLLT can significantly enhance tissue repair and decrease inflammation.7,18,19 Moreover, other investigations have shown that it enhances sperm characteristics, such as motility and survival.7 As indicated by our previous research, using LLLT may help increase the number of germ cells and sperm produced.20 The mechanism by which lasers affect chemical pathways, however, remains unknown. Therefore, the current study has focused on investigating how laser therapy might affect the expression of genes involved in spermatogenesis as well as the activation of molecular pathways.
The process of spermatogenesis plays a crucial role in the transmission of genetic information to the next generation. Understanding this process is crucial for progress in the treatment of human infertility. Spermatogenesis relies on the coordinated activity of multiple geneswhose encoded proteins are involved in the proper testis development, accurate germ cell differentiation, and correct sperm formation and motility. Studying expression patterns in these proteins within different germ cells at various stages can help better understand human spermatogenesis.21 Deleted in azoospermia-like (DAZL) and (DEAD-box helicase 4, DDX4; VASA) genes encode germ cell-specific RNA-binding proteins essential for germ cell development and sperm formation.22,23 Synaptonemal complex protein 3 (SYCP3) is a structural component of the synaptonemal complex required for meiotic chromosome pairing, with mutations linked to azoospermia.24,25 TRIM36 (Haprin) functions in acrosome reaction and cytoskeletal organization,26 while PRM (protamine) is crucial for chromatin condensation in spermatozoa.27 The cation channel of sperm (CatSper) mediates calcium influx and controls sperm motility,28 and G protein-coupled receptor 125 (GPR125; ADGRA3) is a marker of spermatogonial stem cells (SSCs).29 In addition, Acrosin (ACR) is a sperm-specific protease that enables zona pellucida penetration during fertilization.30 These genes ensure proper testicular development, germ cell differentiation, and normal sperm function.
This research focuses on the induction effects of LLLT on spermatogonial stimulation and the underlying mechanisms involved. The study seeks to determine the effects of low-power laser on spermatogenesis in mouse models that present azoospermia. Additionally, the expression levels of key genes associated with differentiation and reproductive processes were analyzed using real-time polymerase chain reaction (RT-PCR).
Methods
Preparation and maintenance of laboratory animals
Adult male NMRI mice, 8–9 weeks old and weighing 30–35 g, were used in this study. All mice were obtained from Drug Applied Research Center and then were kept under controlled temperature conditions (22 ± 2 °C) with a 12-h light/dark cycle and had free access to food and water. All laboratory procedures on mice were conducted in accordance with the guidelines approved by the National Committee for Ethics in Biomedical Research (IRB approval No. IR.IAU.TABRIZ.REC.1403.166).31
In vivo study design
Twenty-four mice were randomly assigned to three groups, each containing 8 animals. The negative control group remained healthy without any treatment. The positive control group included mice in which azoospermia was induced by a single intraperitoneal injection of busulfan (B-2635; Sigma-Aldrich, USA) at 30 mg/kg, dissolved in 0.2 ml of a 1:1 mixture of dimethyl sulfoxide and distilled water, following previously described protocols.32 The experimental group also received busulfan in the same manner and, after 30 days, underwent LLLT using a pulsed diode laser treatment with an energy density of 8 J/cm2, a working frequency of 5 Hz, and a wavelength of 808 nm, applied on a 0.25 cm2 area. The laser was transcutaneously applied to the testicular area for 80 seconds per session, every other day for 3 weeks. During the entire laser treatment, the temperature of the irradiated area was monitored with a thermometer and maintained at 37 °C.
In vitro study design
For the in vitro experiment, testicular tissue was collected from 6 busulfan-treated azoospermic mice, and SSCs were isolated and cultured under standard conditions (37 °C, 5% CO2). The cultured cells were divided into two groups; the control group was not exposed to a laser, and the other received continuous-wave diode laser treatment with an 808 nm wavelength and an energy density of 4 J/cm2 for 53 seconds on a 2.5 cm2 cross-section, with four repetitions every other day. During the laser treatment, the temperature of the irradiated area was monitored using a thermometer and maintained at 37 °C.
Histomorphometric examination of testicular tissue
Following the experimental period's completion, the mice studied in vivo were euthanized for the histomorphometric analysis of the testicular tissue. The testicular tissue was extracted and prepared for staining through tissue sectioning. These sections were then immersed in 10% formalin and stained using hematoxylin-eosin. The morphometric analysis involved the comparison of the epithelium thickness of the seminiferous tubules, the diameter of the seminiferous tubules, and the thickness of the interstitial tissue.
Examining testicular tissue for spermatogenesis
Three fundamental factors, including the differentiation index, the renewability index, and the spermatogenesis index, were analyzed to assess spermatogenesis in testicular tissue. The tubular differentiation index reflects the vitality and differentiation of SSCs, and the renewability index determines the ratio of active spermatogonia to inactive spermatogonia in seminiferous tubules. Further, the spermatogenesis index indicates the proportion of seminiferous tubules containing sperm to those without sperm. In this regard, the cross-sectional areas of the seminiferous tubules were counted and calculated.
Isolation and culture of spermatogonial cells of mouse testicular tissue
Testicular tissue obtained from normal and azoospermic mice was placed in Petri dishes filled with Hank’s balanced salt solution supplemented with 1% antibiotic penicillin (5000 U/mL; Sigma-Aldrich, USA) and streptomycin (5000 mg/mL; Sigma-Aldrich, USA). The capsule surrounding the testicle and epididymis was removed and discarded, after which the testicular tissue was finely chopped using a scalpel. These tissue fragments were then exposed to an enzymatic digestion solution containing ethylene diamine tetra acetic acid -Trypsin (1 mg/mL; Sigma-Aldrich, USA) and type IV collagenase enzyme (1 mg/mL; Sigma-Aldrich, USA). Next, the mixture of cells and enzymes was placed in an incubator set at 37 °C with 5% carbon dioxide for 15 minutes. Throughout this period, pipetting was performed multiple times to ensure proper separation of tissue fragments. Subsequently, the mixture was vortexed for 5 minutes and returned to the incubator for an additional 20 minutes to complete the enzymatic digestion and cell separation process. To counteract the enzyme activity, a complete culture medium consisting of DMEM-F12 (Gibco, USA), supplemented with 10% fetal bovine serum (FBS; Gibco, USA) and 1% Pen/Strep antibiotic, was added, followed by centrifugation at 1,500 rpm for 5 minutes at 4 °C. The supernatant was removed, and the remaining cell pellet was treated with the DMEM-F12 complete culture medium containing 20% FBS and 1% Pen/Strep antibiotic. Cell viability was assessed using a 0.04% trypan blue solution (Sigma-Aldrich, USA) and a hemocytometer. Finally, the cell suspension was transferred to cell culture flasks and maintained at 37°C with 96% humidity and 5% CO2.33
Spermatogonial cells colonization
The colonies of SSCs were observed in testicular tissue culture after 10 days. After mechanical separation, spermatogonial cells were transferred to another flask containing the DMEM-F12 complete culture medium with 20% FBS and 1% Pen/Strep antibiotic. After the first day of cell culture, the culture flask was microscopically examined every day, and the culture medium of the flask was replaced with a new culture medium every 3 days.
Extraction and RT-PCR
Total RNA was isolated from testicular tissue cells and spermatogonial cells using a Trizol-based kit (Sinaclon, Iran) following the manufacturer’s protocol for in vivo and in vitro conditions. Subsequently, the purified RNA was quantified using nanodrop to assess protein or mineral contamination, and its quality was evaluated through electrophoresis on 1% agarose gel. Next, complementary DNA was synthesized using the company’s synthesis kit (Yektatajhiz, Iran). The quantitative changes in the messenger RNA expression of GPR125, VASA, DAZL, SYCP3, ACR, PRM, and TRIM36 genes were analyzed using the RT-PCR technique and the StepOne device (Applied Biosystems, USA). The RT-PCR was performed in 3 replicates using the SYBER Green kit (Ampliqon, Denmark). The master mix of the RT-PCR was prepared with a final volume of 25 μl, consisting of 12.5 μl of SYBR Green, 0.5 μl of the forward primer, 0.5 μl of the reverse primer, 10.5 μl of deoxyribonuclease-free water, and 1 μl of the complementary DNA template added to each microtube. Additionally, the GAPDH gene was utilized as a housekeeping gene. The primers were designed using Oligo Analyzer and Oligo7 software (Table 1), and specific primers for each gene were synthesized by Metabion Company (Germany).
Table 1.
The sequence and properties of applied primers
|
Gene
|
Accession Number
|
Primer Sequence
|
Length
|
TM
|
|
ADGRA3 (GPR125)
|
NM_133911.1 |
F: ACCTGACGAACAACCGAATAG
R: CCGCAACGAGCCAAGATAA |
137 bp |
57°C |
|
DAZL
|
NM_001277863.2 |
F: ATGACGTGGATGTGCAGAAG
R: GAACTGTGGTGGAGGAGGA |
152 bp |
57°C |
|
VASA (DDX4)
|
NM_010029.2 |
F: GGAGATGAAGATTGGGAAGCA
R: TGATGAAGCTGGAGTCCTGT |
126 bp |
58°C |
|
SYCP3
|
NM_011517.2 |
F: GGTTTCCTCAGATGCTTCG
R: AGCCTTTTCATCAGCAACATC |
209 bp |
57°C |
|
PRM
|
NM_013637.5 |
F: CAGCAAAAGCAGGAGCAGA
R: TTCAAGATGTGGCGAGATGC |
212 bp |
58°C |
|
ACR
|
NM_001277247.2 |
F: AATACCCCACACCTGCTACG
R: CCCTGCACACACATTAGTCG |
166 bp |
57°C |
|
HAPRIN (TRIM36)
|
NM_001170855.1 |
F: CAACAGTCCCTATGCGTTCC
R: TTCAGGTTCAGCAAGAGGTG |
159 bp |
57°C |
Statistical analysis
The quantitative data were statistically analyzed using GraphPad Prism 8 software. The results are presented as means ± standard deviations. Statistical significance was assessed through the Student’s t-test and one-way analysis of variance (ANOVA), with differences deemed significant at P < 0.05. The fold-change expression of target genes after treatment was calculated by the ΔΔCT method.
Results
Morphological findings of testicular tissue in in vivo studied groups
In the examination of testicular morphology in the azoospermic group, there was a significant reduction in the quantity of germinal epithelial cells within the seminiferous tubules. A minimal population of spermatogenic cells, predominantly spermatogonia, was detected. Concurrently, there was an expansion of interstitial tissue and a reduction in the diameter of the seminiferous tubules. The majority of seminiferous tubules exhibited the absence of spermatogenic cells within their walls (Fig. 1).
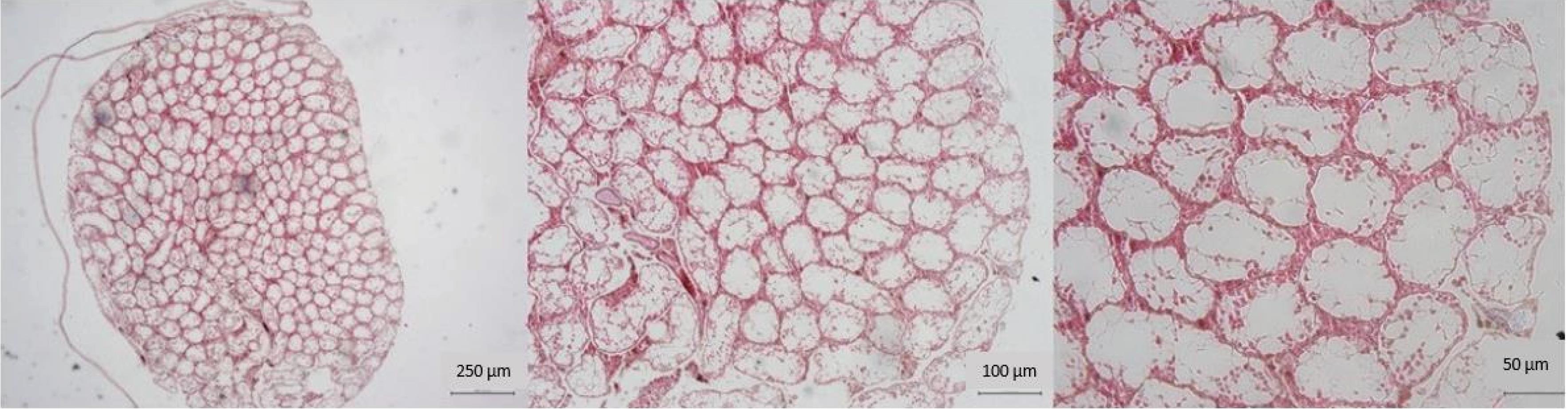
Fig. 1.
Microscopic view of testicular tissue in the positive control azoospermia group. Note. There is a reduction in the diameter of the seminiferous tubules. In addition, there is a severe reduction in the thickness of the tubular epithelium. Spermatogenesis cells are found in very small numbers in the walls of the tubes (40x, 100x, and 200x magnification, hematoxylin-eosin staining).
.
Microscopic view of testicular tissue in the positive control azoospermia group. Note. There is a reduction in the diameter of the seminiferous tubules. In addition, there is a severe reduction in the thickness of the tubular epithelium. Spermatogenesis cells are found in very small numbers in the walls of the tubes (40x, 100x, and 200x magnification, hematoxylin-eosin staining).
In the group that received a dose of 8 J/cm2, the repair process of the testicular tissue was faster. In addition, there was a notable increase in the diameter of the seminiferous tubules and the thickness of their epithelial lining, signifying a positive healing response to the pathological lesions in the testicle. Additionally, there was a rise in the quantity of germ cells within the tubular walls (Fig. 2).
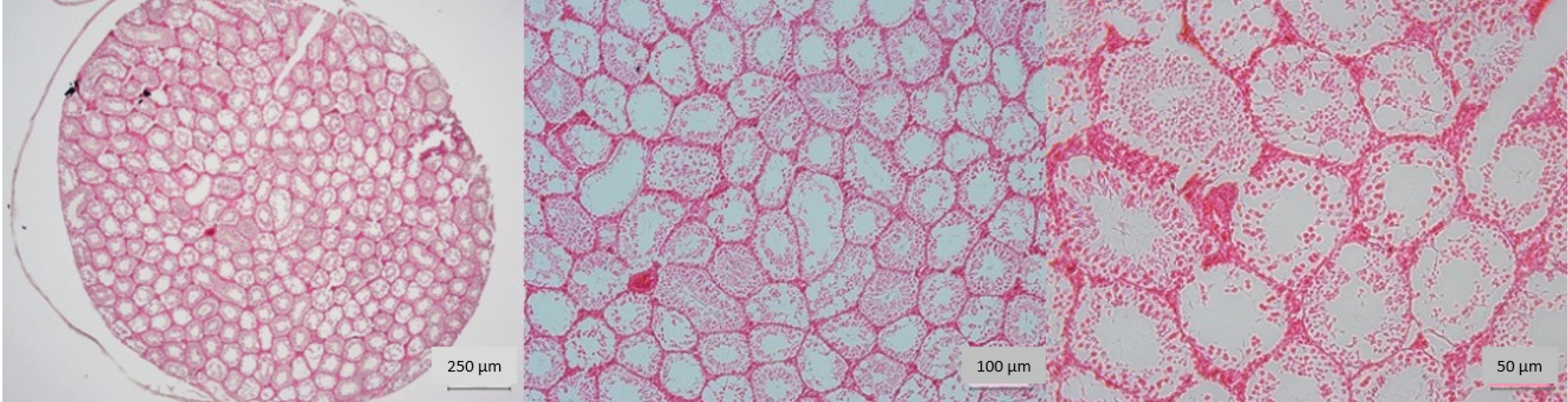
Fig. 2.
Microscopic view of testicular tissue in the azoospermia group with 8J/cm2 energy density irradiation. Note. Testicular tissue has repaired, and there is an increase in the diameter of the seminiferous tubule. Moreover, the tubular epithelium has been repaired. The number of spermatogenesis cells has increased sharply (40x, 100x, and 200x magnification, hematoxylin-eosin staining).
.
Microscopic view of testicular tissue in the azoospermia group with 8J/cm2 energy density irradiation. Note. Testicular tissue has repaired, and there is an increase in the diameter of the seminiferous tubule. Moreover, the tubular epithelium has been repaired. The number of spermatogenesis cells has increased sharply (40x, 100x, and 200x magnification, hematoxylin-eosin staining).
Testicular spermatogenesis results
The analysis results of the tubular differentiation index coefficient indicated a significant increase in the group that received 8 J/cm2 laser energy compared to the positive control group (P < 0.05). No significant differences were found between the groups in terms of the spermatogenesis index coefficient (P ˃ 0.05). In the analysis of the renewability index, a significant increase was observed in the group exposed to 8 J/cm2 laser energy compared to the positive control group (P < 0.05).
Gene expression analysis of the in vivo study
The expression levels of VASA and DAZL, genes associated with spermatogenesis, were evaluated to confirm the morphological impact of LLLP on azoospermia. The results of our study confirmed a significant reduction in VASA expression (P < 0.05) observed four weeks following the administration of busulfan (Fig. 3A). On the other hand, VASA expression was upregulated after the irradiation of the diode laser at 8J/cm2 (Fig. 3B).
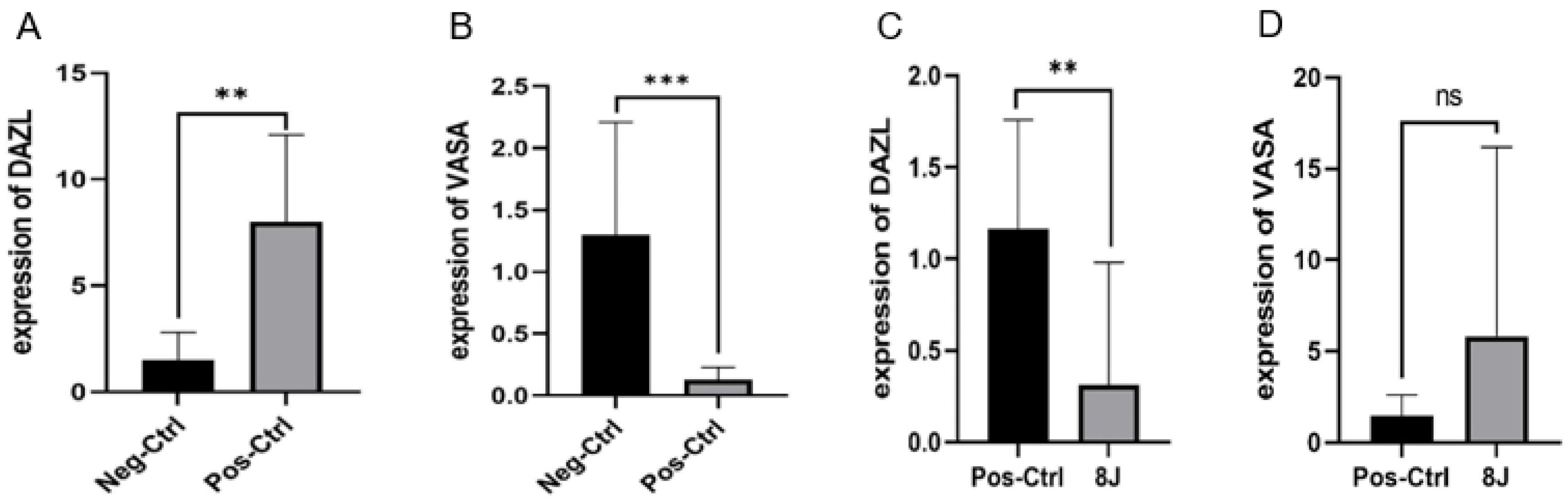
Fig. 3.
Comparison of the expression ratios of DAZL and VASA between the two groups using a t-test under in vivo study conditions, with 8 mice per group. Note. To make the expression differences between the groups visible and understandable, the graphs are based on the t-test. (A) and (B) Comparison of expression levels in the positive control versus negative control groups for DAZL and VASA. In this comparison, DAZL expression showed a significant increase (P < 0.05) after busulfan injection in the positive control group, while VASA expression displayed a significant decrease (P < 0.05). (C) and (D) Comparison of expression levels in the positive control versus laser treatment groups for DAZL and VASA. Following laser irradiation, DAZL expression demonstrated a noticeable decrease (P < 0.05), whereas VASA expression depicted a slight, non-significant increase. ns indicates non-significance. ** and ***represent significant differences.
.
Comparison of the expression ratios of DAZL and VASA between the two groups using a t-test under in vivo study conditions, with 8 mice per group. Note. To make the expression differences between the groups visible and understandable, the graphs are based on the t-test. (A) and (B) Comparison of expression levels in the positive control versus negative control groups for DAZL and VASA. In this comparison, DAZL expression showed a significant increase (P < 0.05) after busulfan injection in the positive control group, while VASA expression displayed a significant decrease (P < 0.05). (C) and (D) Comparison of expression levels in the positive control versus laser treatment groups for DAZL and VASA. Following laser irradiation, DAZL expression demonstrated a noticeable decrease (P < 0.05), whereas VASA expression depicted a slight, non-significant increase. ns indicates non-significance. ** and ***represent significant differences.
It was observed that the expression of DAZL was significantly increased in the azoospermic model (P< 0.05, Fig. 3D). However, a significant down-regulation was noted after the application of laser treatment (P < 0.05). Table 2 presents the fold-change results of the comparison between the positive and negative control groups, as well as the comparison of the treatment group with the positive control group.
Table 2.
Fold changes between the studied groups
|
Gene
|
Ctrl + /Ctrl-
|
8J/Ctrl +
|
|
Fold change
|
P
value
|
Fold change
|
P
value
|
|
DAZL
|
+ 6.8824 |
0.0087 |
-14.6498 |
0.0017 |
|
VASA
|
-11.3858 |
0.0055 |
+ 1.75828 |
0.5205 |
Note. Ctrl + : Control positive; Ctrl-: Control negative; 8J: Laser density at 8J/cm2.
Culture and colony formation of spermatogonial stem cells (in vitro)
SSCs were isolated from the testicular tissue of adult mice, which were subsequently cultured and expanded. The testicular tissue underwent a two-day incubation period, during which the cells adhered to the flask’s surface and began to proliferate. The spermatogonial cells are characterized by their large size and smooth, regular morphology, often exhibiting 2–3 eccentric nuclei (Fig. 4A).
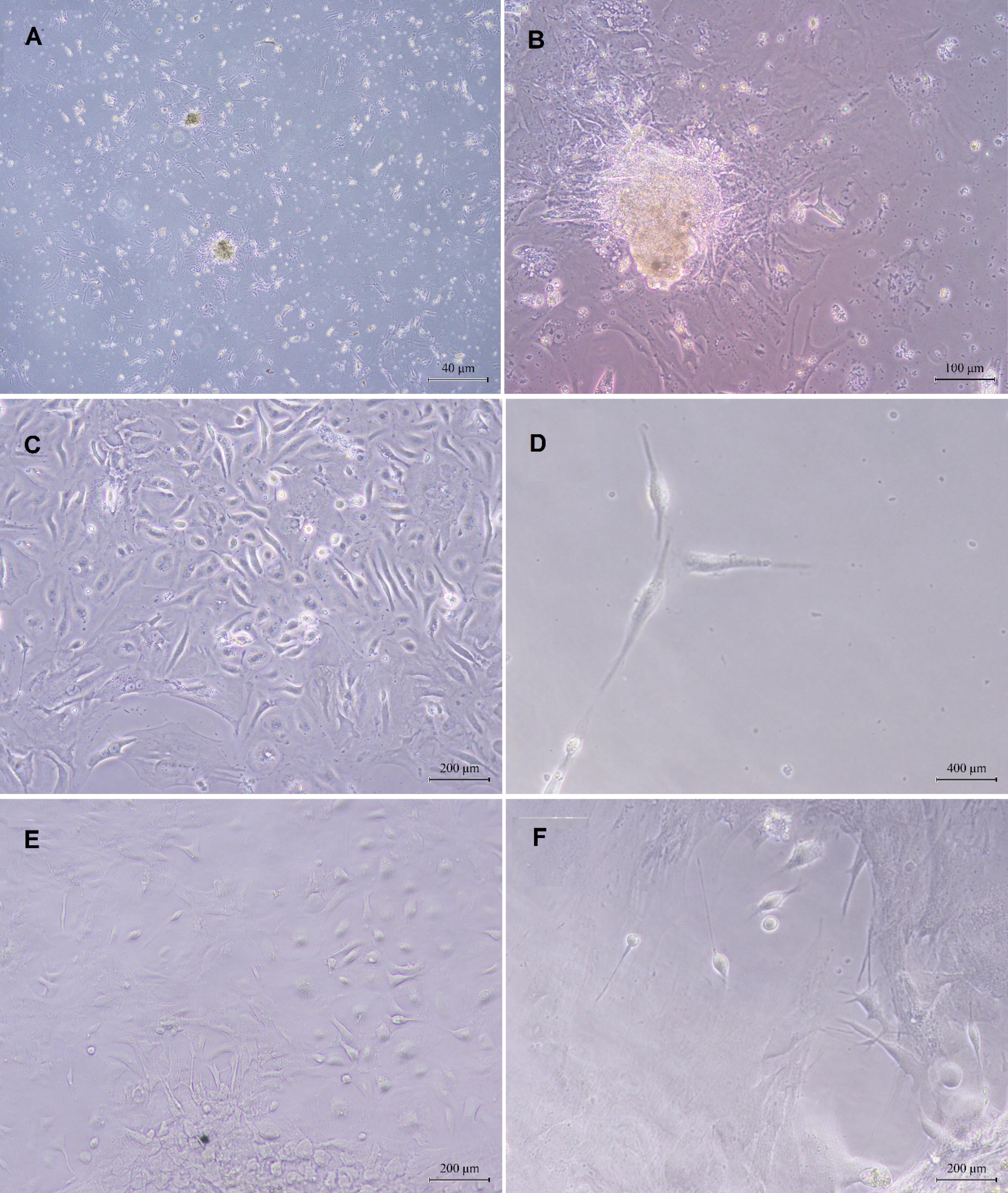
Fig. 4.
Microscopic images obtained from the culture of spermatogonial cells and their differentiation toward mature sperm production: (A) Testicular tissue culture and colony growth of spermatogonial stem cells two days after primary culture, (B) Increased density and area of spermatogonial colonies seven days after primary culture, (C, D) Microscopic image of cell differentiation after two weeks of laser irradiation, and (E, F) Microscopic image of cell differentiation into sperm cells after three weeks of laser irradiation.
.
Microscopic images obtained from the culture of spermatogonial cells and their differentiation toward mature sperm production: (A) Testicular tissue culture and colony growth of spermatogonial stem cells two days after primary culture, (B) Increased density and area of spermatogonial colonies seven days after primary culture, (C, D) Microscopic image of cell differentiation after two weeks of laser irradiation, and (E, F) Microscopic image of cell differentiation into sperm cells after three weeks of laser irradiation.
The cultivation of spermatogonial cells in a culture medium initiates a process of growth and division, resulting in the formation of cell clusters. Over a period of 5–7 days, these clusters develop into dense, typically spherical masses known as colonies. Initially, these colonies can be observed at low cell densities. After two weeks, the cells achieve a significantly increased density in terms of the number of colonies and the area they occupy (Fig. 4B).
Induction of spermatogonial stem cells by laser (in vitro)
The testicular tissue of adult mice that were treated with busulfan was utilized for the production of SSCs. After being cultivated in 24-well plates, the SSCs were subjected to an 808 nm wavelength, 75 mW of power, 53 seconds of irradiation time, and 4 J/cm2 of energy. One side of the cells swelled and acquired a semi-sperm shape after a week, and the appendages on both sides of the cell appeared more elongated due to the laser’s impact on the cells.
The morphology of SSCs continued to change toward sperm production after two weeks; in the second week, there was a rise in resemblance and a denser nuclear area (Figs. 4C and 4D).
The cells changed in form, became more similar to each other, and resembled sperm in the third week after induction. Their cytoplasm lengthened, nuclei became more compressed, the head took on an oval shape, and they developed a long appendage resembling a tail (Figs. 4E and 4F).
Gene expression analysis of the in vitro study
Gene expression analysis was performed at two specific stages: after busulfan injection and after laser treatment.
The expression level of the marker genes involved in the spermatogenesis process was investigated at two-week and three-week intervals after the culture of spermatogonial cells and exposure to an energy dose of 4 J/cm2 in 4 repetitions with an interval of one day.
After busulfan injection, the gene expression level was evaluated in the positive control group two weeks later. The expression of each gene significantly decreased compared to the healthy negative control group, as indicated by RT-PCR data analysis results. DAZL and VASA genes in this group exhibited a slight change, which was not statistically significant, while the SYCP3 gene showed the most significant alteration. Moreover, there was a notable increase (P < 0.05) in the GPR125 gene expression levels compared to the negative control group. Details regarding fold change and P-value are provided in Table 3. Graphs illustrating the comparison between each gene in the positive and negative control groups are depicted in Fig. 5.
Table 3.
Comparison of the expression level of the busulfan positive control group with the negative control group
|
Gene
|
Ctrl-/Ctrl +
|
|
Fold change
|
P
value
|
|
DAZL
|
-1.2022 |
0.7430 |
|
VASA
|
-1.1019 |
0.0761 |
|
GPR125
|
2.2501 |
0.0098 |
|
SYCP3
|
-7.2358 |
0.0459 |
|
ACR
|
-2.5053 |
0.0443 |
|
TRIM36
|
-3.0194 |
0.0010 |
|
PRM
|
-2.2345 |
0.0351 |
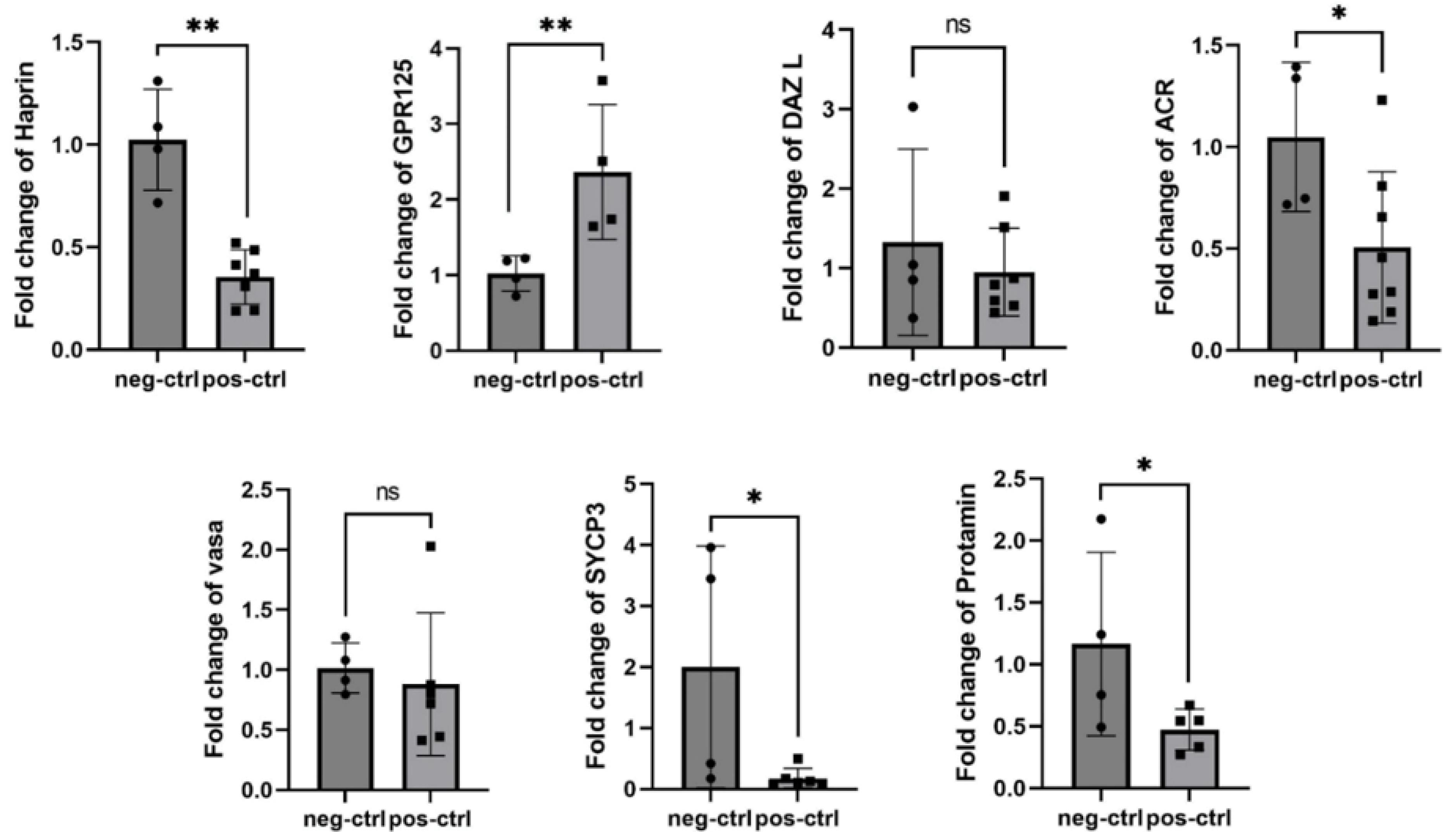
Fig. 5.
Comparison of the expression levels of seven genes (GPR125, VASA, DAZL, SYCP3, ACR, PRM, and TRIM36) between the two control groups (negative and positive) using a t-test under in vitro study conditions in 6 test mice. Note. All genes, except for GPR125, showed decreased expression in the positive control group after busulfan injection. Although these decreases were statistically significant (P < 0.05) for all genes, VASA and DAZL did not display significant differences. ns represents non-significant difference. * and **represent significant differences.
.
Comparison of the expression levels of seven genes (GPR125, VASA, DAZL, SYCP3, ACR, PRM, and TRIM36) between the two control groups (negative and positive) using a t-test under in vitro study conditions in 6 test mice. Note. All genes, except for GPR125, showed decreased expression in the positive control group after busulfan injection. Although these decreases were statistically significant (P < 0.05) for all genes, VASA and DAZL did not display significant differences. ns represents non-significant difference. * and **represent significant differences.
Most genes analyzed in this study demonstrated significant increases following irradiation with an energy dose of 4 J/cm2 during laser treatment. Two treatment groups, the 3-week and 2-week radiation groups, were compared, and the results revealed that genes such as ACR, PRM, and Sycp3 exhibited higher expression levels after two weeks of radiation than after three weeks. Conversely, DAZL, GPR125, TRIM36, and VASA displayed increased expression after three weeks of radiation exposure. The data related to fold change and P value are presented in Table 4, along with graphs depicting the comparison between each gene’s positive and negative control groups (Fig. 6).
Table 4.
Comparison of the expression of the 4 J/cm2 treatment group for 2 weeks and 3 weeks
|
Gene
|
Ctrl + /4J-2W
|
Ctrl + /4J-3W
|
|
Fold change
|
P
value
|
Fold change
|
P
value
|
|
DAZL
|
+ 1.5910 |
0.0432 |
+ 2.3254 |
0.0127 |
|
VASA
|
+ 1.6330 |
0.0915 |
+ 1.9976 |
0.0420 |
|
GPR125
|
-2.7368 |
0.3429 |
+ 2.3254 |
0.1328 |
|
SYCP3
|
+ 3.9449 |
0.0346 |
+ 2.9173 |
0.0980 |
|
ACR
|
+ 2.1983 |
0.0242 |
+ 2.0244 |
0.2491 |
|
TRIM36
|
-1.2989 |
0.9027 |
+ 2.8664 |
0.0254 |
|
PRM
|
+ 2.0326 |
0.0086 |
+ 1.9690 |
0.0079 |
Note. Ctrl + : Control positive; 4J-2w: Laser density at 4J/cm2 after 2 weeks; 4J-3w: Laser density at 4J/cm2 after 3 weeks.
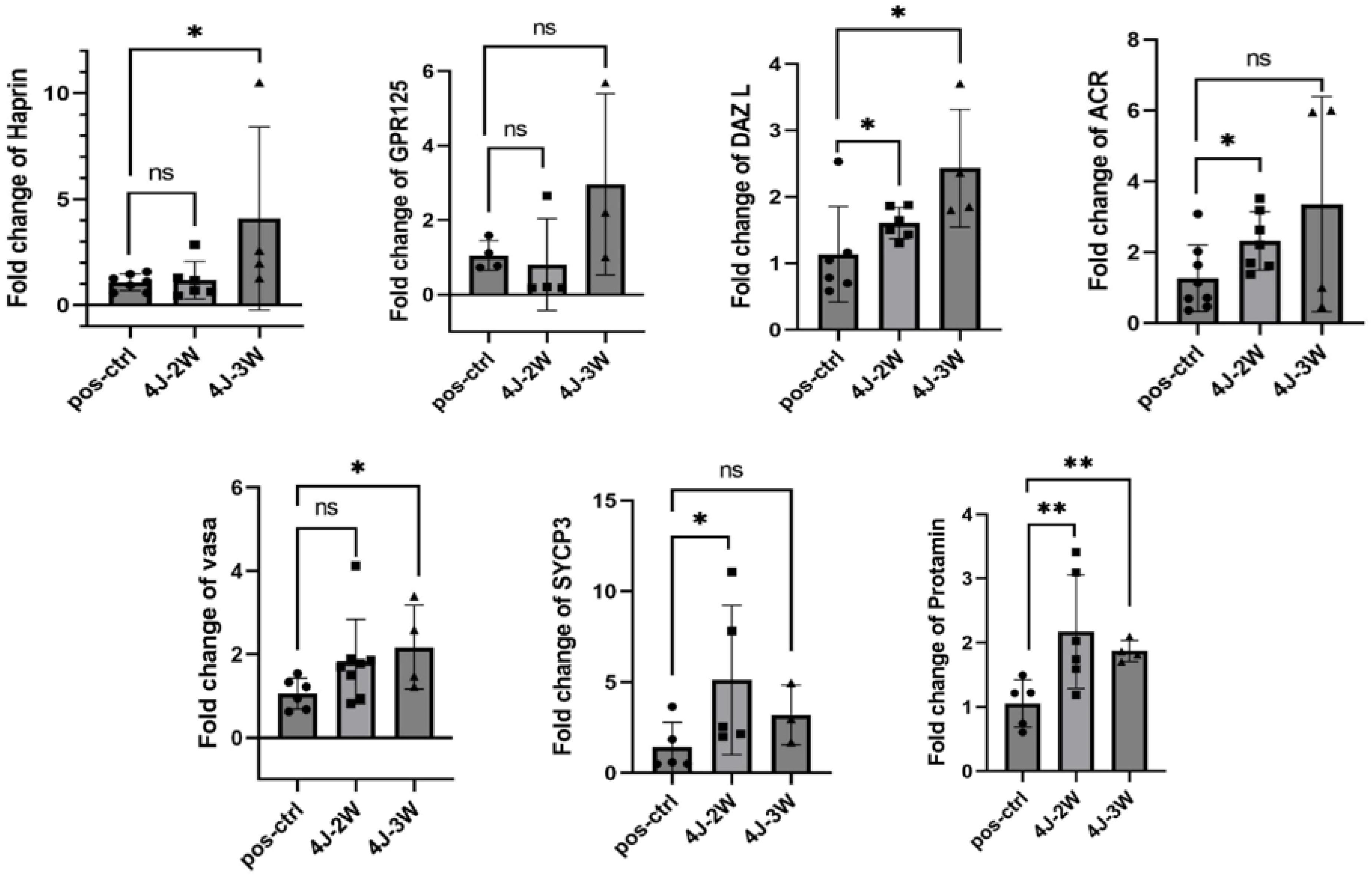
Fig. 6.
Comparison of the expression levels of seven studied genes (GPR125, VASA, DAZL, SYCP3, ACR, PRM, and TRIM36) in three positive control groups and under laser treatment for two weeks and three weeks. Note. ANOVA: One-way analysis of variance.This comparison was conducted using one-way ANOVA under in vitro study conditions. In the second week following laser irradiation, an increase was observed in the expression of the studied genes, with most changes reaching statistical significance (P < 0.05); however, a slight, non-significant decrease was noted for GPR125 and TRIM36. In the third week, the expression of all genes increased, although the increase was not statistically significant for some genes.ns represents non-significant difference. * and ** represent significant differences.
.
Comparison of the expression levels of seven studied genes (GPR125, VASA, DAZL, SYCP3, ACR, PRM, and TRIM36) in three positive control groups and under laser treatment for two weeks and three weeks. Note. ANOVA: One-way analysis of variance.This comparison was conducted using one-way ANOVA under in vitro study conditions. In the second week following laser irradiation, an increase was observed in the expression of the studied genes, with most changes reaching statistical significance (P < 0.05); however, a slight, non-significant decrease was noted for GPR125 and TRIM36. In the third week, the expression of all genes increased, although the increase was not statistically significant for some genes.ns represents non-significant difference. * and ** represent significant differences.
Discussion
Despite recent studies demonstrating the effect of low-power lasers on the normal growth of the testis and the treatment of male infertility, a thorough understanding of how laser affects molecular pathways and the mechanism underlying its effect on normal spermatogenesis remains unknown. This study evaluated the impact of laser light on the sperm production process in the infertile azoospermia mouse model. The results indicated that, at an appropriate dosage, laser irradiation can be a successful treatment for azoospermia caused by busulfan. These findings are supported by concurrent in vivo and in vitro observations, suggesting photobiomodulation's dose-dependent, stage-specific effect on spermatogenesis.
In-vivo, exposure to 8 J/cm2 laser irradiation produced accelerated testicular healing, with an increased number of germ cells within the testicular wall and enhancements in the diameter and thickness of the tubular epithelium. In vitro, laser treatment at 4 J/cm2 induced a morphological shift in cultured SSCs from a fibroblast like to a sperm like phenotype during weeks 2–3, indicating the initiation of spermatogenic progression under photobiomodulation. Overall, these structural and morphological changes align with subsequent gene expression findings, supporting the coordinated laser-induced promotion of spermatogenesis.
To study the impact of lasers, the expression levels of some important genes were assessed, including those involved in sperm production and differentiation both in vitro and in vivo (i.e., VASA, DAZ L, GPR125, SYCP3, ACR, TRIM36, and PRM). Busulfan disrupts the expression of these genes, which is consistent with its known antifertility effects. In contrast, laser irradiation reversed these disruptions, with dose- and context-dependent trajectories.
Busulfan is known to induce azoospermia by germ cell loss and suppression of germ cell gene expression, as well as apoptosis promotion and differentiation inhibition within the spermatogenic lineage. Our data align with this understanding and extend it by demonstrating that photobiomodulation can counteract busulfan’s antifertility effects, at least at the examined doses. The observed upregulation of germ cell- and maturation-associated genes under laser treatment is in line with the broader literature, reporting the regenerative effects of low-power lasers on testicular tissue and germ cell markers in related models. The previous study’s morphological and histomorphological results confirmed that busulfan, at an effective dose of 30 mg, causes azoospermia by reducing the number of sperm cells34,35; however, this study’s examination and comparison of the expression level of marker genes in the control group and the busulfan experimental group in both in vivo and in vitro conditions indicated that busulfan injection significantly reduces the expression of testicular specific genes. This alkylating drug inhibits cell division by binding to the DNA chain. It further causes damage to germ cells and disrupts the process of spermatogenesis, resulting in a decrease in the weight of the testicles.34-37 In vitro experiments showed that the expression of genes associated with spermiogenesis, such as SYCP3, ACR, TRIM36, and PRM, decreased significantly. Similarly, it was found that the expression of the VASA gene, which usually begins with the onset of spermatogenesis, also decreased significantly in-vivo. However, three weeks after the administration of busulfan, the expression of GPR125 and DAZL genes showed a considerable increase. In our in vitro study, SSCs were isolated from mouse testicular tissue treated with busulfan and then cultured. Despite the inhibitory effect of busulfan, we anticipated an increase in the expression of the GPR125 gene compared to the control group, which consists of various differentiated cells. GPR125 is a marker of undifferentiated spermatogonial germ cells and is believed to denote the initial cells entering the spermatogenesis stage.38-40 Furthermore, it was revealed that, in contrast to what was anticipated, the expression of DAZL, a hallmark of spermatogonia and primary germ cells, was elevated in-vivo.41,42 The histological inspection of the testis in the azoospermic group in the previous work demonstrated the presence of several spermatogenic cell types, primarily spermatogonial cells, even after a thorough evaluation of the seminiferous tubules following busulfan injection.43 Therefore, it makes sense that DAZL gene expression would still be high in azoospermic settings. Even though the in vitro study showed a reduction in this gene’s expression, it was not statistically significant, and more research is required in this regard.
Busulfan generally destroys germ cell lines by promoting apoptosis and reducing cell division. It also stops the few spermatogonial cells that remain from differentiating into mature sperm and from advancing the spermatogenesis process.44 Studies have been performed thus far to reduce and even reverse busulfan’s antifertility impact.45,46 These investigations demonstrate that the antifertility effect of busulfan is curable and offers a means of utilizing less complex and risky therapy techniques, including photobiomodulation.
In recent years, many infertility-related illnesses have been treated using the photobiomodulation approach. Our findings conform to those of previous studies17,47 about the use of laser therapy to treat infertility. Low-power lasers have been shown to have a regenerative impact on testicular tissue. According to our results, using an energy density of 4 J/cm2 for two or three weeks significantly boosted marker gene expression and initiated the spermatogenesis process in vitro study circumstances.
Our findings confirmed that the expression of these genes significantly increased two weeks after the laser irradiation, and they had somewhat maintained their increasing trend in the third week. These markers of germ cells include GPR125, VASA, and DAZL genes, which are expressed in the early stages of the development and differentiation of spermatogonial cells,48 and the four genes SYCP3, ACR, TRIM36, and PRM, which are markers of differentiated spermatogonial and sperm maturation.
The GPR125 gene expression represented a slight declining trend after two weeks of laser irradiation, which was statistically insignificant. Three weeks later, however, its expression began to be upregulated with an energy density of 4 J/cm2; therefore, the population of SSCs increased. Some studies reported that the expression of this gene is unnecessary for normal spermatogenesis, but the lack of GPR125 gene expression prevents the proper maturation of sperm and fertility. According to research performed by Nybo et al, 55% of mouse models with no expression of the GPR125 gene were infertile from puberty, even though they had normal spermatogenesis and several epididymal sperm. These mice also had transiently enlarged epididymis, decreased alpha estrogen receptors, and increased production of estradiol. The GPR125 gene is essential for the growth of the male reproductive system because it’s functions in tandem with estrogen signaling. When this gene is not expressed, hormonal imbalances occur after puberty, obstructing the distal reproductive system and resulting in the buildup of fluid and sperm in the epididymis.49 This work increased the expression of the GPR125 gene in the culture of spermatogonial cells isolated from the testis treated with busulfan. This upward trend continued after the laser’s effect, suggesting that the GPR125 gene is active from the start of cell development. Fertility and sperm maturation depend on the spermatogonia foundation.
Two and three weeks of laser radiation at a dosage of 4 J/cm2 resulted in a slow but significant rise in the level of DAZL and VASA gene expression. These particular germ cell genes have protective roles in the development and fertility of both humans and animals.50 The DAZL gene product can be regarded as a marker gene in the male gametogenesis process since it causes the germ cell to progress toward meiosis and produce a haploid cell. Numerous abnormalities in mouse spermatogenesis, including mitotic dysregulation and meiotic arrest, can result from the loss of DAZL expression and function.51 Additionally, a study has shown that by inhibiting the pluripotency program and hindering somatic cell differentiation, primordial germ cells can reduce the risk of testicular teratoma formation by suppressing pluripotency factors through DAZL in vitro.52 On the other hand, since the VASA gene generates a protein that enhances sperm motility and germ cell differentiation in the testis, its presence is crucial during the entire process of spermatogenesis.53,54 Mice that had the VASA gene deleted were rendered infertile despite the creation of primary germ cells as a result of aberrant zygote spermatocyte growth, colonization, and death.55 Some studies56,57 indicate that these two genes co-express each other. However, there is no proof that the expression of one of these genes leads to the upregulation of another. Although the enhanced expression of DAZL and VASA was observed in the current investigation in vitro up to the third week, the in vivo trial with the 8 J/cm2 radiation dose showed the opposite amount of expression. Following laser irradiation, DAZL displayed a considerable decrease compared to the busulfan group, despite VASA gene expression rising dramatically. This phenomenon may be explained by several theories, such as variations in how cells react to laser treatment, the impact of microenvironmental elements, and the influence of systemic components that are absent in vitro but present in vivo.
DAZL and VASA do not appear to work in concert, but their overexpression raises the percentage of ACR positive cells, suggesting that both proteins may serve as indicators for ACR gene expression.58,59 According to this study, the elevated levels of DAZL and VASA are linked to elevated expression of the ACR gene. Under the low-power laser irradiation of 4 J/cm2, this gene's expression gradually increased significantly compared to the busulfan positive control group. In a different recent work, human Wharton’s jelly mesenchyme cells were exposed to LED radiation at a wavelength of 625 nm and an energy of 1.9 J/cm2, which led to the differentiation of the cells toward gametogenesis in vitro and an increase in the expression of DAZL and VASA genes, along with ACR.59 It can be concluded that while the laser itself can induce the expression of the ACR gene by triggering a cascade of events or factors, the high expression of the two pre-meiotic genes (DAZL and VASA) also contributes to the effective expression of the ACR gene.
SYCP360 is another gene linked to the expression of DAZL and VASA. The second week of laser irradiation in this investigation was when the expression of this gene increased the most. This gene is exclusively expressed in testicular tissue, and its protein induces the formation of a synaptonemal complex between homologous chromosomes during the meiotic prophase of spermatogenesis. As a result, it is commonly referred to as a meiosis marker.61,62 Male advanced zygote spermatocytes may undergo apoptosis if SYCP3 is absent.63 This gene’s expression starts in primary spermatocytes, which is consistent with its function in promoting meiosis.64 In the second week of laser irradiation, these genes had the highest expression, indicating the density of spermatogonial cells and spermatocytes in the environment. This was observed in this study by analyzing the expression level of the SYCP3 gene and comparing it with the level of expression of markers of the early stages of sperm development.
Although they began to increase from the second week, the expression of TRIM36 and PRM genes, which are thought to be markers of sperm maturation and are expressed in spermatid and spermatozoa,65-67 showed less expression until the second week in comparison to pre-puberty genes. The expression of these two genes rose during the third week of laser irradiation, and spermatogonial cell cultures were examined under a microscope during this same period, revealing the presence of mature sperm with proper morphology. The TRIM36 gene is highly expressed in the testis and haploid germ cells. It generates a ubiquitinating protein that, by breaking down target proteins with the aid of the proteasome complex, assists in chromosomal movement during the cell cycle. This protein is essential to the sperm acrosome region’s proper functioning.68,69 Thus, as spermatogenesis progresses, its expression progressively rises. While normal spermatogenesis was previously observed in a homozygous heparin-deficient mouse model, sperm quality, morphology, and motility were lower than in wild-type animals,65 indicating the importance of this gene expression during proper spermatogenesis. Additionally, the PRM gene encodes chromatin-bound proteins that help the sperm head’s DNA condense and organize.66 Histone retention and an increase in reactive oxygen species are the two main causes of DNA damage caused by low protamine gene expression, leading to less healthy sperm and an increase in sperm motility issues.67
Based on in vitro results, laser irradiation at 4 J/cm2 led to a significant upregulation of germ cell gene expression over 2–3 weeks, indicating the initiation and progression of spermatogenesis-like processes in the SSC cultures. Conversely, in vivo model results at 8 J/cm2 revealed structural and cellular improvements in the testicular tissue and gene expression changes that support the restoration of spermatogenic programs, though temporal and magnitude differences between in vitro and in vivo contexts reflect systemic influences, the microenvironment, and the complex regulation in a living organism. These data suggest that noninvasive tissue photobiomodulation could mitigate busulfan-induced infertility by promoting germ cell gene expression and activity, facilitating meiosis and supporting germ cell maturation. Potential mechanisms may include shifts in cellular metabolism and mitochondrial activity, signaling pathways governing differentiation, and changes in the testicular niche that support spermatogenesis. However, functional assessments (e.g., sperm counts, motility, and fertilization capacity), long-term safety evaluations, and genetic integrity are needed before clinical translation. The strengths of this study were, a comprehensive design across in vitro and in vivo models, dose-consistent effects, multi-gene expression profiling across stages of spermatogenesis, and demonstration of a plausible, dose-dependent therapeutic window exist. On the other hand, the limitations of this study included gene-expression–based evidence without full functional sperm outputs, lack of longer-term safety data, and the caution required when extrapolating differences between in vitro and in vivo conditions to humans. Safety considerations involve more precise assessments of genetic integrity and any off-target effects to remain safe prior to any clinical application. Future implications include expanding functional assessments to examine sperm counts, motility, and fertilization capacity, evaluating DNA health after laser treatment, examining a broader range of doses and exposure patterns to define an optimal therapeutic window, investigating molecular pathways and signaling cascades that mediate photobiomodulation effects in germ cells and the testicular niche, and validating findings in larger animal models and, where appropriate and safe, in human tissue or cells under rigorous ethical and safety controls. In general, noninvasive laser therapy, at appropriate doses, is capable of mitigating busulfan-induced infertility by promoting germ cell gene expression and spermatogenesis progression in vitro and by improving testicular tissue structure in-vivo. These findings support the potential of photobiomodulation as a therapeutic avenue for male infertility while emphasizing the need for replication across models, a deeper understanding of underlying mechanisms, thorough safety evaluations, and demonstration of functional fertility outcomes before clinical application.
Conclusion
The process of producing sperm from the mouse models of azoospermia was studied in this study, both in vitro and in vivo. The in vitro results revealed that laser therapy with an energy density of 8J/cm2 could enhance germ cells. Sperm production also had benefits. Conversely, the energy density of 4J/cm2, which was employed to investigate the level of gene expression associated with the spermatogenesis process in a laboratory setting, demonstrated the efficacy of this radiation dosage in vitro. As a result, our findings confirmed that increasing the generation of natural sperm and treating infertility can be achieved by the employment of a low-power diode laser with a wavelength of 808 nm in both laboratory and in vivo settings.
Research Highlights
What is the current knowledge?
-
Azoospermia, characterized by the absence of sperm in the ejaculate, affects approximately 1% of the male population and significantly contributes to male infertility.
-
Low-power laser therapy has been explored for its potential therapeutic effects on various biological processes, including tissue regeneration and cellular function.
What is new here?
-
The findings of this study demonstrated that low-power 808 nm diode laser therapy can enhance sperm production in azoospermic mice, indicating a novel therapeutic approach for male infertility.
-
The research identified specific gene expression changes, particularly the upregulation of VASA, associated with improved spermatogenesis following laser treatment.
-
Both in vivo and in vitro models were utilized to validate the efficacy of laser therapy, providing a comprehensive understanding of its impact on male fertility.
Competing Interests
None.
Data Availability Statement
The raw data and analytical methods utilized in this study are available upon reasonable request from the corresponding author.
Ethical Approval
This study was performed in line with the principles of Islamic Azad University of Medical Sciences, Tabriz (IRB approval No. IR.IAU.TABRIZ.REC.1403.166).
Acknowledgements
The authors acknowledge Biophotonics and Biotechnology Research Center, Islamic Azad University of Tabriz, for its scientific support.
References
- World Health Organization (WHO). Infertility Prevalence Estimates, 1990-2021. WHO; 2023.
- Kumar N, Singh AK. Trends of male factor infertility, an important cause of infertility: A review of literature. J Hum Reprod Sci 2015; 8:191-6. doi: 10.4103/0974-1208.170370 [Crossref] [ Google Scholar]
- Rezaei F, Bayat M, Nazarian H, Aliaghaei A, Abaszadeh HA, Naserzadeh P. Photobiomodulation therapy improves spermatogenesis in busulfan-induced infertile mouse. Reprod Sci 2021; 28:2789-2798. doi: 10.1007/s43032-021-00557-8 [Crossref] [ Google Scholar]
- He Z, Kokkinaki M, Jiang J, Zeng W, Dobrinski I, Dym M. Isolation of human male germ-line stem cells using enzymatic digestion and magnetic-activated cell sorting. Methods Mol Biol 2012; 825:45-57. doi: 10.1007/978-1-61779-436-0_4 [Crossref] [ Google Scholar]
- Esteves SC, Ramasamy R, Colpi GM, Carvalho JF, Schlegel PN. Sperm retrieval rates by micro-TESE versus conventional TESE in men with non-obstructive azoospermia—the assumption of independence in effect sizes might lead to misleading conclusions. Hum Reprod Update 2020; 26:603-5. doi: 10.1093/humupd/dmaa006 [Crossref] [ Google Scholar]
- Borovets SY, Moskvin SV. Laser therapy techniques for the treatment of male infertility. Herald Urology 2017; 5:49-61. doi: 10.21886/2308-6424-2017-5-3-49-61 [Crossref] [ Google Scholar]
- Ziaeipour S, Norouzian M, Abbaszadeh HA, Aliaghaei A, Nazarian H, Karamian A. Photobiomodulation therapy reverses spermatogenesis arrest in hyperthermia-induced azoospermia mouse model. Lasers Med Sci 2023; 38:114. doi: 10.1007/s10103-023-03780-8 [Crossref] [ Google Scholar]
- Deihimi M, Azornia M, Takzare N, Rajab M, Hasanzadeh GH. Effect of red and infrared spectrum low level of laser rays on Rat Seminiferous tubules. J Gorgan Univ Med Sci 2010; 12:10-17. [ Google Scholar]
- Dompe C, Moncrieff L, Matys J, Grzech-Leśniak K, Kocherova I, Bryja A. Photobiomodulation-Underlying Mechanism and Clinical Applications. J Clin Med 2020; 9:1724. doi: 10.3390/jcm9061724 [Crossref] [ Google Scholar]
- Tsuka Y, Kunimatsu R, Gunji H, Abe T, Medina CC, Nakajima K. Examination of the Effect of the Combined Use of Nd: YAG Laser Irradiation and Mechanical Force Loading on Bone Metabolism Using Cultured Human Osteoblasts. J Lasers Med Sci 2020; 11:138-143. doi: 10.34172/jlms.2020.24 [Crossref] [ Google Scholar]
- Zare F, Moradi A, Fallahnezhad S, Ghoreishi SK, Amini A, Chien S. Photobiomodulation with 630 plus 810nm wavelengths induce more in vitro cell viability of human adipose stem cells than human bone marrow-derived stem cells. J Photochem Photobiol Biol 2019; 201:111658. doi: 10.1016/j.jphotobiol.2019.111658 [Crossref] [ Google Scholar]
- Garrido PR, Pedroni ACF, Cury DP. Effects of photobiomodulation therapy on the extracellular matrix of human dental pulp cell sheets. J Photochem Photobiol B 2019; 194:149-157. doi: 10.1016/j.jphotobiol.2019.03.017 [Crossref] [ Google Scholar]
- Ohsugi Y, Niimi H, Shimohira T, Hatasa M, Katagiri S, Aoki A. In Vitro Cytological Responses against Laser Photobiomodulation for Periodontal Regeneration. Int J Mol Sci 2020; 21:9002. doi: 10.3390/ijms21239002 [Crossref] [ Google Scholar]
- Siqueira AF, Maria FS, Mendes CM, Hamilton TR, Dalmazzo A, Dreyer TR. Effects of photobiomodulation therapy (PBMT) on bovine sperm function. Lasers Med Sci 2016; 31:1245-1250. doi: 10.1007/s10103-016-1966-z [Crossref] [ Google Scholar]
- Iaffaldano N, Meluzzi A, Manchisi A, Passarella S. Improvement of stored turkey semen quality as a result of He-Ne laser irradiation. Anim Reprod Sci 2005; 85:317-325. doi: 10.1016/j.anireprosci.2004.04.043 [Crossref] [ Google Scholar]
- Corral-Baqués MI, Rigau T, Rivera M, Rodríguez JE, Rigau J. Effect of 655-nm diode laser on dog sperm motility. Lasers Med Sci 2005; 20:28-34. doi: 10.1007/s10103-005-0332-3 [Crossref] [ Google Scholar]
- Gabel CP, Carroll J, Harrison K. Sperm motility is enhanced by Low Level Laser and Light Emitting Diode photobiomodulation with a dose-dependent response and differential effects in fresh and frozen samples. Laser Ther 2018; 27:131-136. doi: 10.5978/islsm.18-or-13 [Crossref] [ Google Scholar]
- Preece D, Chow KW, Gomez-Godinez V, Gustafson K, Esener S, Ravida N. Red light improves spermatozoa motility and does not induce oxidative DNA damage. Nature 2017; 7:19. doi: 10.1038/srep46480 [Crossref] [ Google Scholar]
- Deihimi M, Azornia M, Takzare N, Rajab M. Hasanzadeh Effect of red and infrared spectrum low level of laser rays on rat Seminiferous tubules. J Gorgan Uni Med Sci 2010; 12:1017. [ Google Scholar]
- Tajalli H, Maleki M, Safavi E, Shahi R, Firoozi F, Akbarpour Z. The effects of low-power laser on the promotion of spermatogenesis in a mouse model of azoospermia (in-vivo). IJBBE 2022; 2:15-30. doi: 10.30495/ijbbe.2022.698214 [Crossref] [ Google Scholar]
- Amirian M, Azizi H, Hashemi Karoii D, Skutella T. VASA protein and gene expression analysis of human non-obstructive azoospermia and normal by immunohistochemistry, immunocytochemistry, and bioinformatics analysis. Sci Rep 2022; 12:17259. doi: 10.1038/s41598-022-22137-9 [Crossref] [ Google Scholar]
- Li H, Liang Z, Yang J, Wang D, Wang H, Zhu Zhu, M M. DAZL is a master translational regulator of murine spermatogenesis. Nat Sci Rev 2019; 6:455-468. doi: 10.1093/nsr/nwy163 [Crossref] [ Google Scholar]
- Azizi H, Ranjbar M, Rahaiee S, Govahi M, Skutella T. Investigation of VASA Gene and Protein Expression in Neonate and Adult Testicular Germ Cells in Mice In Vivo and In Vitro. Cell J 2022; 22:171-177. doi: 10.22074/cellj.2020.6619 [Crossref] [ Google Scholar]
- Aarabi M, Soltanghoraee H, Aarabi M, Behjati Ardakani R, Amirjannati N, Ghaffari M, et al. Expression of Synaptonemal Complex Protein 3 (SYCP3) mRNA in the testis: a molecular marker for spermatogenesis in azoospermic men. J Reprod Infertil 2006. 6: 495-504.
- Tsutsumi M, Kogo H, Kowa-Sugiyama H, Inagaki H, Ohye T, Kurahashi H. Characterization of a Novel Mouse Gene Encoding an SYCP3-Like Protein That Relocalizes from the XY Body to the Nucleolus During Prophase of Male Meiosis I. Biol Reprod 2011; 85:165-171. doi: 10.1095/biolreprod.110.087270 [Crossref] [ Google Scholar]
- Barros CP, Derks MFL, Mohr J, Wood BJ, Crooijmans RPMA, Megens HJ. A new haplotype-resolved turkey genome to enable turkey genetics and genomics research. Gigascience 2022; 12:giad051. doi: 10.1093/gigascience/giad051 [Crossref] [ Google Scholar]
- Akmal M, Aulanni'am A, Aris Widodo M, Sumitro S, Purnomo B, Widodo N. The important role of protamine in spermatogenesis and quality of sperm: A mini review. Asian Pac J Reprod 2016; 5:357-360. doi: 10.1016/j.apjr.2016.07.013 [Crossref] [ Google Scholar]
- Jin ZR, Fang D, Liu BH, Cai J, Tang WH, Jiang H. Roles of CatSper channels in the pathogenesis of asthenozoospermia and the therapeutic effects of acupuncture-like treatment on asthenozoospermia. Theranostics 2021; 11:2822-2844. doi: 10.7150/thno.51869 [Crossref] [ Google Scholar]
- Izadyar F, Wong J, Maki C, Pacchiarotti J, Ramos T, Howerton K. Identification and characterization of repopulating spermatogonia stem cells from the adult human testis. Hum Reprod 2011; 26:1296-1306. doi: 10.1093/humrep/der026 [Crossref] [ Google Scholar]
- Tranter R, Read JA, Jones R, Brady RL. Effector sites in the three-dimensional structure of mammalian sperm beta-acrosin. Structure 2000; 8:1179-1188. doi: 10.1016/s0969-2126(00)00523-2 [Crossref] [ Google Scholar]
- Guide for the care and use of laboratory animals. 2020. https://ethics.research.ac.ir/EthicsProposalView.php?&code=IR.IAU.TABRIZ.REC.1404.353.
- Wang DZ, Zhou XH, Yuan YL, Zheng XM. Optimal dose of busulfan for depleting testicular germ cells of recipient mice before spermatogonial transplantation. Asian J Androl 2010; 12:263-70. doi: 10.1038/aja.2009.67 [Crossref] [ Google Scholar]
- Bahrami M, Rahimi-Feyli P, Moghaddam AA. Effect of vitamin C on colony formation of ovine spermatogonial stem cells in vitro. SJKU 2017; 22:32-43. [ Google Scholar]
- Zohni K, Zhang X, Tan SL, Chan P, Nagano MC. the efficiency of male fertility restoration is dependent on the recovery kinetics of spermatogonial stem cells after cytotoxic treatment with busulfan in mice. Hum Reprod 2012; 27:44-53. doi: 10.1093/humrep/der357 [Crossref] [ Google Scholar]
- Chegini R, Soleimani P, Sadeghi M, Mohammad Yosef R, Zafari F. Investigating the effect of fennel and cinnamon combined extract on spermatogenesis and testis tissues in busulfan induced infertile rats. J Appl Biotechnol Rep 2019; 6:96-100. doi: 10.29252/JABR.06.03.03 [Crossref] [ Google Scholar]
- Allameh F, Razzaghi M, Hosseini S, Barati M, Razzaghi Z, Salehi S. The effect of laser acupuncture on semen parameters in infertile men with oligospermia: a randomized clinical trial. J Lasers Med Sci 2021; 12:e84. doi: 10.34172/jlms.2021.84 [Crossref] [ Google Scholar]
- Qu N, Itoh M, Sakabe K. Effects of Chemotherapy and Radiotherapy on Spermatogenesis: The Role of Testicular Immunology. Int J Mol Sci 2019; 20:957. doi: 10.3390/ijms20040957 [Crossref] [ Google Scholar]
- Seandel M, James D, Shmelkov SV, Falciatori I. Generation of functional multipotent adult stem cells from GPR125+ germline progenitors. Nature 2007; 449:346-350. doi: 10.1038/nature06129 [Crossref] [ Google Scholar]
- Golestaneh N, Kokkinaki M, Pant D, Jiang J. Pluripotent stem cells drived from adult human testes. Stem Cells Dev 2009; 18:1115-1126. doi: 10.1089/scd.2008.0347 [Crossref] [ Google Scholar]
- Izadyar F, Wong J, Maki C, Pacchiarotti J, Ramos T, Howerton K. Identification and characterization of repopulating spermatogonia stem cells from the adult human testis. Hum Reprod 2011; 26:1296-1306. doi: 10.1093/humrep/der026 [Crossref] [ Google Scholar]
-
Kee K, Angles VT, Flores M, Nguyen HN, Rejio Pera RA. Human DAZL, DAZ and BOULE genes modulate primordial germ-cell and haploid gamete formation. Nature 2009; 12: 462: 222-225. doi: 10.1038/nature08562.
- Reijo RA, Dorfman DM, Slee R, Renshaw AA, Loughlin KR, Cooke H. DAZ family proteins exist throughout male germ cell development and transit from nucleus to cytoplasm at meiosis in humans and mice. Biol Reprod 2000; 63:1490-1496. doi: 10.1095/biolreprod63.5.1490 [Crossref] [ Google Scholar]
- Tajalli H, Maleki M, Safavi E, Shahi R, Firoozi F, Akbarpour Z. The effects of low-power laser on the promotion of spermatogenesis in a mouse model of azoospermia (in-vivo). IJBBE 2022; 2:15-30. doi: 10.30495/ijbbe.2022.698214 [Crossref] [ Google Scholar]
- Choi YJ, Ok DW, Kwon DN, Chung JI, Kim HC, Yeo SM. Murine male germ cell apoptosis induced by busulfan treatment correlates with loss of c-kit-expression in a Fas/FasL-and p53 independent manner. FEBS Lett 2004; 575:41-51. doi: 10.1016/j.febslet.2004.08.034 [Crossref] [ Google Scholar]
- Tamadon A, Mehrabani D, Rahmanifar F, Jahromi AR, Panahi M, Zare S. Induction of spermatogenesis by bone marrow-derived mesenchymal stem cells in busulfan-induced azoospermia in hamster. Int J Stem Cells 2015; 8:134. doi: 10.15283/ijsc.2015.8.2.134 [Crossref] [ Google Scholar]
- Deihimi M, Azornia M, Takzare N, Rajab M, Hasanzadeh GH. Effect of red and infrared spectrum low level of laser rays on Rat Seminiferous tubules. J Gorgan Univ Med Sci 2010; 12:10-17. [ Google Scholar]
-
Dreyer T, Siqueira A. Biochemical and topological analysis of bovine sperm cells induced by low power laser irradiation. European Conference on Biomedical Optics, Optical Society of America; 2011. doi: 10.1364/ecbo.2011.80920v.
- Shlush E, Maghen L, Swanson S, Kenigsberg S, Moskovtsev S, Barretto T. In vitro generation of Sertoli-like and haploid spermatid-like cells from human umbilical cord perivascular cells. Stem Cell Res Ther 2017; 8:37. doi: 10.1186/s13287-017-0491-8 [Crossref] [ Google Scholar]
- Nybo ML, Kvam JM, Nielsen JE, Frederiksen H, Spiess K, Jensen KHR. Loss of Adgra3 causes obstructive azoospermia with high penetrance in male mice. FASEB J 2023; 37:e22781. doi: 10.1096/fj.202200762RR [Crossref] [ Google Scholar]
- Zhu W, Wang T, Zhao Ch, Wang D, Zhang X, Zhang H. Evolutionary conservation and divergence of Vasa, Dazl and Nanos1 during embryogenesis and gametogenesis in dark sleeper [Odontobutis potamophila]. Gene 2018; 672:21-33. doi: 10.1016/j.gene.2018.06.016 [Crossref] [ Google Scholar]
- Li T, Wang X, Zhang H, Chen H, Liu N, Xue R. Gene expression patterns and protein cellular localization suggest a novel role for DAZL in developing Tibetan sheep testes. Gene 2022; 731:144335. doi: 10.1016/j.gene.2020.144335 [Crossref] [ Google Scholar]
- Chen HH, Welling M, Bloch DB, Muñoz J, Mientjes E, Chen X, Tramp C. DAZL Limits Pluripotency, Differentiation, and Apoptosis in Developing Primordial Germ Cells. Stem Cell Rep 2014; 3:892-904. doi: 10.1016/j.stemcr.2014.09.003 [Crossref] [ Google Scholar]
- Castrillon DH, Quade BJ, Wan TY, Quigley C, Crum CP. The human VASA gene is specifically expressed in germ cell lineage. Proc Natl Acad Sci USA 2000; 97:9585-9590. doi: 10.1073/pnas.160274797 [Crossref] [ Google Scholar]
- Li HJ, Yu N, Zhang XY, Jin W, Li HZ. Spermatozoal protein profiles in male infertility with asthenozoospermia. Chin Md J 2010; 123:2879-2882. doi: 10.3760/cma.j.issn.0366-6999.2010.20.024 [Crossref] [ Google Scholar]
- Amirian M, Azizi H, Hashemi Karoii D, Skutella T. VASA protein and gene expression analysis of human non-obstructive azoospermia and normal by immunohistochemistry, immunocytochemistry, and bioinformatics analysis. Sci Rep 2022; 12:17259. doi: 10.1038/s41598-022-22137-9 [Crossref] [ Google Scholar]
- Kito G, Tanaka H, Soh T, Yamauchi N, Hattori MA. The putative promoters of germ cell-specific genes and Nanog are hypomethylated in chicken sperm. J Reprod Dev 2014; 60:224-229. doi: 10.1262/jrd.2014-010 [Crossref] [ Google Scholar]
- Luo Y, Xie L, Mohsin A, Ahmed W, Xu C, Peng Y. Efficient generation of male germ-like cells derived during co-culturing of adipose-derived mesenchymal stem cells with Sertoli cells under retinoic acid and testosterone induction. Stem Cell Res Ther 2019; 10:91. doi: 10.1186/s13287-019-1181-5 [Crossref] [ Google Scholar]
- Torres-Fernández LA, Emich J, Port Y, Mitschka S, Wöste M, Schneider S. TRIM71 deficiency causes germ cell loss during mouse embryogenesis and is associated with human male infertility. Front Cell Dev Biol 2021; 9:970. doi: 10.3389/fcell.2021.658966 [Crossref] [ Google Scholar]
- Azizi H, Niazi Tabar A, Skutella T. Successful transplantation of spermatogonial stem cells into the seminiferous tubules of busulfan-treated mice. Reprod Health 2021; 18:1-9. doi: 10.1186/s12978-021-01242-4 [Crossref] [ Google Scholar]
- Rosario R, Adams IR, Anderson Ra. Is there a role for DAZL in human female fertility?. Mol Hum Reprod 2016; 22:377-383. doi: 10.1093/molehr/gaw024 [Crossref] [ Google Scholar]
- Myamoto Myamoto, T. T., Hasuike Hasuike, S. S., Yogev Yogev, L L. Azoospermia in patients heterozygous from a mutation in SYCP3. Lancet 2003; 362:1714-1719. doi: 10.1016/s0140-6736(03)14845-3 [Crossref] [ Google Scholar]
- Martinez-Garay I, Jablonka S, Sutajova M, Steuernagel P, Gal A, Kutsche K. A new gene family [FAM9] of low-copy repeats in Xp223 expressed exclusively in testis. Genomics 2002; 80:259-267. doi: 10.1006/geno.2002.6834 [Crossref] [ Google Scholar]
- Yuan L, Liu JG, Zhao J, Brundell E, Daneholt B, Hoog C. The murine SCP3 gene is required for synaptonemal complex assembly, chromosome synapsis, and male fertility. Mol Cell 2000; 5:73-83. doi: 10.1016/s1097-2765(00)80404-9 [Crossref] [ Google Scholar]
- Aarabi M, Modarressi MH, Soltanghoraee H, Behjati R, Amirjannati N, Akhondi Akhondi, MM MM. Testicular expression of synaptonemal complex protein 3 (SYCP3) messenger ribonucleic acid in 110 patients with nonobstructive azoospermia. Fertil Steril 2006; 86:325-331. doi: 10.1016/j.fertnstert.2005.12.070 [Crossref] [ Google Scholar]
- Aoki Y, Tsujimura A, Kaseda K, Okabe M, Tokuhiro K, Ohta T. Haprin-deficient spermatozoa are incapable of in vitro fertilization. Mol Reprod Dev 2022; 87:534-541. doi: 10.1002/mrd.23344 [Crossref] [ Google Scholar]
- Kanippayoor RL, Alpern JH, Moehring AJ. Protamines and spermatogenesis in Drosophila and Homo sapiens: A comparative analysis. Spermatogenesis 2013; 3:e24376. doi: 10.4161/spmg.24376 [Crossref] [ Google Scholar]
- Akmal M. Aulanni'am A, Aris Widodo M, Sumitro S, Purnomo B, Widodo N The important role of protamine in spermatogenesis and quality of sperm: A mini review. Asian Pac J Reprod 2016; 5:357-360. doi: 10.1016/j.apjr.2016.07.013 [Crossref] [ Google Scholar]
- Balint I, Muller A, Nagy A, Kovacs G. Cloning and characterization of the RBCC728/TRIM36 zinc-binding protein from the tumor suppressor gene region at chromosome 5q223. Gene 2004; 332:45-50. doi: 10.1016/j.gene.2004.02.045 [Crossref] [ Google Scholar]
- Kitamura K, Nishimura H, Nishimune Y, Tanaka H. Identification of human HAPRIN potentially involved in the acrosome reaction. J Androl 2005; 26, 4:511-518. doi: 10.2164/jandrol.041 [Crossref] [ Google Scholar]