Bioimpacts. 15:31430.
doi: 10.34172/bi.31430
Review
Ambivalent roles of miRNAs in cancer development via modulating tumor-associated innate immune cells
Bahar Naseri Writing – original draft, 1, # 
Amirhossein Mardi Writing – original draft, 1, 2, #
Najibeh Shekari Writing – original draft, 2
Neda Shajari Writing – original draft, 3, 1
Samin Abdolzadeh Writing – review & editing, 1
Hossein Khorramdelazad Writing – review & editing, 4
Amirhossein Hatami-Sadr Writing – review & editing, 1
Milad Taghizadeh Anvar Visualization, 5
Mohammad Reza Javan Conceptualization, 6
Amirhossein Heibatollahi Visualization, 1
Javad Masoumi Conceptualization, 1
Farid Ghorbaninezhad Supervision, 7, *
Behzad Baradaran Supervision, 1, 2, *
Author information:
1Immunology Research Center, Tabriz University of Medical Sciences, Tabriz, Iran
2Department of Immunology, Faculty of Medicine, Tabriz University of Medical Sciences, Tabriz, Iran
3Cancer Immunology and Immunotherapy Research Center, Ardabil University of Medical Sciences, Ardabil, Iran
4Department of Immunology, School of Medicine, Rafsanjan University of Medical Sciences, Rafsanjan, Iran
5Department of Immunology, School of Medicine, Shahid Beheshti University of Medical Sciences, Tehran, Iran
6Department of Immunology, Faculty of Medicine, Zabol University of Medical Sciences, Zabol, Iran
7Student Research Committee, Department of Immunology, School of Medicine, Shahid Beheshti University of Medical Sciences, Tehran, Iran
#These authors contributed equally to this study.
Abstract
The tumor microenvironment (TME), comprising malignant and non-transformed cells like immune cells, endothelial cells, and cancer-associated fibroblasts, significantly affects tumor growth and progression. Tumor cells manipulate the TME by releasing chemokines and inhibitory cytokines, reprogramming surrounding cells to support their survival and evade immune detection. Innate immune cells within the TME play dual roles, either promoting or inhibiting tumor progression, impacting immunotherapy outcomes. Recent studies highlight the influence of innate immune cells in shaping the TME and the pivotal role of tumor-derived microRNAs (miRNAs) in modulating these cells. miRNAs regulate gene expression and enhance tumor immune evasion, angiogenesis, drug resistance, and invasion. Their tumor-specific expression patterns suggest potential as biomarkers and therapeutic targets. This study focuses on how miRNAs affect innate immune cells like macrophages, dendritic cells, myeloid-derived suppressor cells, and natural killer cells, contributing to immunosuppressive or immunogenic environments. Understanding miRNA-mediated interactions between cancer and immune cells opens new possibilities for improving targeted immunotherapy and advancing cancer treatments.
Keywords: Cancer, miRNA, Innate immune system, TME, Biomarker, Immunomodulation
Copyright and License Information
© 2025 The Author(s).
This work is published by BioImpacts as an open access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (
http://creativecommons.org/licenses/by-nc/4.0/). Non-commercial uses of the work are permitted, provided the original work is properly cited.
Funding Statement
This study received no external funding.
Introduction
The most significant attributes of cancer cells include their ability to proliferate uncontrollably, resist apoptosis, exhibit genomic instability, and invade surrounding tissues. In this context, the tumor microenvironment (TME) is vital role in enhancing these capabilities within cancer cells.1 In the cancer literature, different perspectives exist regarding the TME; nevertheless, the most concise elucidation of this term is "the setting that affects the growth, survival, and progression of tumor cells".2 Despite its simplistic definition, TME operates as a sophisticated and well-structured ecosystem. In addition to malignant cells, this milieu also harbors non-transformed cells, e.g., endothelial cells, pericytes, fibroblasts, immune cells, and other cell types that vary depending on the tissue, like neurons and adipocytes.3 Cancer cells' expansion and ability to evade immune surveillance is facilitated by the cellular interactions between cancerous and non-cancerous cells, which serve as the cytological mechanism underlying tumor progression and metastasis.4 For instance, tumor angiogenesis involves a dynamic interplay between endothelial cells and other cells that drive angiogenesis, such as pericytes, vascular smooth muscle cells, macrophages, skeletal muscle cells, and tumor cells. This communication is mediated through various mechanisms, including cell-cell adhesion, junctional complexes formation, and paracrine cytokines and metabolite release5 (Fig. 1). Besides, cancer cells can manipulate their surrounding setting by secreting different chemokines, inhibitory cytokines, and other inhibitory molecules.6 Subsequently, the surrounding cells undergo a reprogramming mechanism, allowing them to assume a pivotal role in the survival and progression of tumors.6 For example, transforming growth factor‐beta (TGF‐β), interleukin (IL)‐6, and platelet‐derived growth factor (PDGF) secreted by tumor cells can activate quiescent fibroblasts and alter them into cancer‐associated fibroblasts (CAFs).7 CAFs are the main contributors to the production of TGF-β. TGF-β inhibits the proliferation of CD4+T lymphocytes by reducing the production of IL-2 and facilitates the differentiation of naive CD4+T lymphocytes into regulatory T cells (Tregs).8 The proliferation of cancer cells is marked by the restructuring of the vasculature and the extracellular matrix (ECM).3 This matrix, predominantly made up of collagen and proteoglycans, provides a scaffold for the cellular microenvironment and contributes to the secretion of various cytokines, chemokines, and other bioactive molecules.9,10 Cancer cells' capability to penetrate the ECM barrier, access the circulatory system, and develop distant metastases plays a vital role in the progression and metastasis of tumors.11 Hence, this intricate network is embedded within an altered, vascularized extracellular matrix, contributing to the organization of tumor settings.3
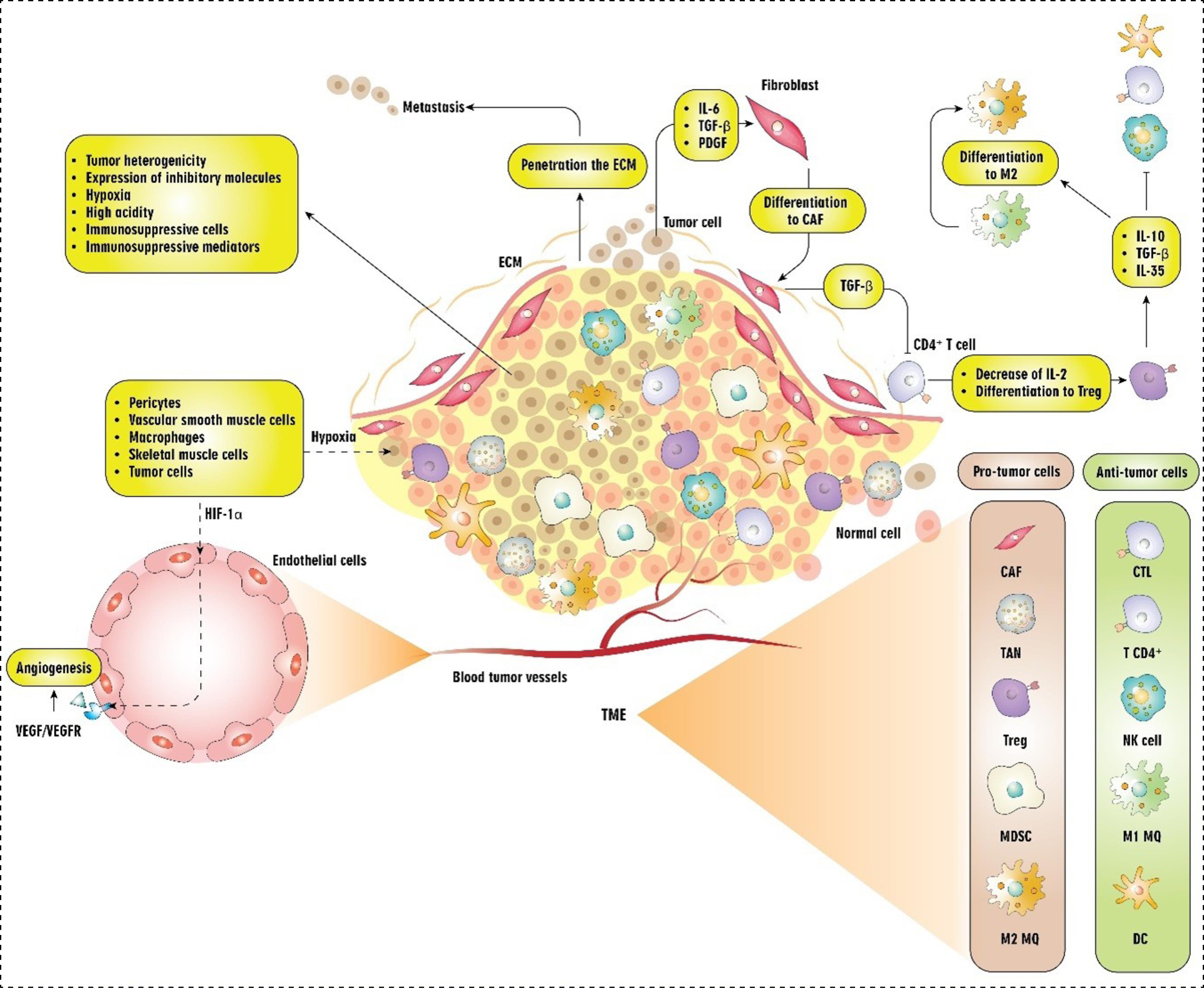
Fig. 1.
Cellular and molecular components of the TME and their interactions. Abbreviations: TME: tumor microenvironment, ECM: extracellular matrix, TAN: tumor-associated neutrophils, NK cell: natural killer cell, Treg: T regulatory cell, DC: dendritic cell, CAF: cancer-associated fibroblast, MDSC: myeloid-derived suppressor cells, CTL: cytotoxic T cells, MQ: macrophage, HIF-1α: Hypoxia-inducible factor-1α, VEGF: Vascular endothelial growth factor, TGF: Transforming growth factor, PDGF: Platelet-derived growth factor.
.
Cellular and molecular components of the TME and their interactions. Abbreviations: TME: tumor microenvironment, ECM: extracellular matrix, TAN: tumor-associated neutrophils, NK cell: natural killer cell, Treg: T regulatory cell, DC: dendritic cell, CAF: cancer-associated fibroblast, MDSC: myeloid-derived suppressor cells, CTL: cytotoxic T cells, MQ: macrophage, HIF-1α: Hypoxia-inducible factor-1α, VEGF: Vascular endothelial growth factor, TGF: Transforming growth factor, PDGF: Platelet-derived growth factor.
Beyond the cellular interactions present within the environment, the significance of tumor heterogeneity cannot be overlooked. Numerous cancer types exhibit considerable heterogeneity in the TME, both spatially and temporally.12 Heterogeneity is described as the diversity observed among cancer cells, both within individual tumors and between different tumors. This diversity includes variations in cellular morphology, transcriptional profiles, metabolic processes, and the capacity for metastasis. The presence of heterogeneity is a common feature in most tumors, presenting considerable challenges within cancer ecosystems. It significantly affects the long-term effectiveness of treatments for solid tumors, leading to resistance, heightened metastatic behavior, and recurrence.12 Due to their genomic instability and capacity for phenotypic variation, cancer cells can quickly modify their behavior to exploit local environmental conditions. A well-documented instance of this is the metabolic symbiosis that takes place between cancer cells in oxygen-deprived regions and those in well-oxygenated parts of tumors.13 Recent advancements in the field have also focused on the TME and the interactions that facilitate the evolution of the tumor ecosystem.12 As previously stated, immune cells are fundamental constituents of the TME.3 Within TME, a heterogeneous population of adaptive and innate immune cells infiltrates, displaying the potential to exert both pro- and anti-tumorigenic impacts.14 The immune landscape surrounding tumors is predominantly characterized by two categories of immune cells: those that promote tumor growth and those that inhibit it. Key pro-tumor immune cells consist of myeloid-derived suppressor cells (MDSCs), Tregs, and M2-type tumor-associated macrophages (TAMs), which collectively enhance tumor development and immune escape mechanisms. In contrast, the anti-tumor immune response is primarily mediated by activated cytotoxic T lymphocytes (CTLs) and natural killer (NK) cells1,15 (Fig. 1). Considering this, the tumor immune microenvironment is a cutting-edge concept that has been associated with the clinical outcomes of cancer patients, predicting and directing their immunotherapy response.16 The tumor immune microenvironment provides valuable insights into the potential trajectory of a patient's immune response. This path of cancer development is shaped by the kinds of immune cells that infiltrate tumors, the presence of inhibitory immune checkpoint molecules on tumor or immune cells, and changes in the TME.17
Scientific reports have highlighted the presence of adaptive and innate immune cells within the tumor milieu.4,18 The main focus of earlier studies has been on the analysis of adaptive immune cells within the context of cancer.19-22 The literature on the TME has expanded, shedding light on the profound influence of innate immune cells.23 It is now apparent that the response of innate immune cells not only indirectly affects the TME by controlling the fate of T cells.24,25 but also plays a critical role in shaping the TME. In this regard, within TME, innate immune cells can play a dual role by either promoting (pro-tumoral innate immune cells).26,27 or inhibiting (anti-tumoral innate immune cells)28,29 tumor progression. Despite the inhibitory role of pro-tumoral innate immune cells in tumor progression, the altering of anti-tumoral innate immune cells by tumor cells within the TME can inhibit their ability to combat tumors by fostering an immunosuppressive environment and metabolic reprogramming.30-32 Additionally, tumor cells can strengthen the ability of pro-tumoral innate immune cells to maintain and perpetuate this immunosuppressive setting.30-32 In line with this, a growing body of scientific studies has shown that cancer-derived microRNAs (miRNAs) have a profound impact on the formation of an immunosuppressive TME.33,34 miRNAs, which are short non-coding RNAs (ncRNAs) consisting of around 22 nucleotides, have a significant impact on the regulation of gene expression. These regulatory molecules modulate gene activity by attaching to the 3’ untranslated region (3’UTR) of target mRNAs, which can lead to reduced gene expression by blocking transcription or protein production.35 miRNAs play a crucial role in modulating the activity of innate immune cells, diminishing the immunogenic potential of cancer cells, and enabling tumor cells to evade detection by the innate immune system.33,34 Besides, the regulation of the TME by miRNAs can lead to changes in tumor angiogenesis,36 drug resistance,37 proliferation of malignant cells,38 and their invasion.39 Hence, the expression of immunity-associated genes in cancer cells and tumor-infiltrating innate immune cells is subject to regulation by miRNAs. Moreover, the expression profiles of miRNAs differ significantly among most tumors, and specific miRNAs can serve as markers to distinguish between various tumor types and their respective stages.40,41 As a result, miRNAs have the potential to be used clinically in the treatment, prognosis, and diagnosis of various cancers (Table 1). Herein, we aim to clarify the impact of tumor cells-related miRNAs on the alteration of the function and characteristics of innate immune cells, including macrophages (MQ), MDSCs, dendritic cells (DCs), NK cells, neutrophils, and γδ T cells toward the modulation of malignancy and the creation of both an immunosuppressive and an immunogenic setting. By underscoring the pivotal role of miRNAs, this study sheds light on the intricate signaling network that exists between cancer cells and innate immune cells, thereby elucidating the crucial involvement of miRNAs in tumor immune evasion. Furthermore, this study highlights the potential avenues for utilizing miRNAs as targets in therapy, thereby augmenting the efficacy of targeted immunotherapy for the treatment of cancer.
Table 1.
Recruiting trials for the clinical applications of miRNAs in various cancers
|
NCT number
|
Cancer type
|
Clinical application
|
Summary of study
|
Enrolled patients
|
| NCT02635087 |
Colonic neoplasms |
Prognostic biomarker |
An observational study determining the use of miRNAs as a tool to predict disease prognosis |
630 |
| NCT03962088 |
Rectal cancer |
A biomarker for monitoring tumor response |
An observational study aiming to evaluate the tumor response to surgery after neoadjuvant chemoradiotherapy of affected patients by examining the expression pattern of miRNAs |
200 |
| NCT04285476 |
Thyroid cancer |
Diagnostic biomarker |
An interventional study evaluating the sensitivity and specificity of miRNAs as a diagnostic tool and risk stratification biomarker |
70 |
| NCT04845425 |
Endometrial cancer |
Diagnostic and prognostic biomarkers |
An observational study aiming to evaluate the miRNA expression pattern to identify biomarkers to better stratify Endometrial cancer patients |
150 |
| NCT05346757 |
CRC |
Screening test |
An interventional study to validate a miRNA-based fecal test for CRC screening |
9670 |
| NCT05918510 |
SCC of the oropharynx, carcinomas of unknown primary sites, high-risk HPV infection |
Diagnostic and prognostic biomarkers |
An observational study evaluating the possibility of miRNAs as a potential diagnostic and prognostic biomarkers |
142 |
| NCT05854030 |
Lung neoplasm |
Prognostic biomarker, a biomarker for monitoring tumor response |
An observational study determining the serum exosomal miRNA in combination with PD-L1 as a biomarker in predicting the efficacy of anti-PD-L1 immunotherapy |
60 |
| NCT04435756 |
Germ cell tumor |
Disease recurrence biomarker, outcome prediction biomarker |
A prospective observational study to evaluate miRNA 371 for outcome prediction in affected patients |
956 |
| NCT06277986 |
GC cachexia |
Diagnostic biomarker |
An observational study evaluating the clinical value of tumor cell-derived exosomal miRNA in the diagnosis of GC cachexia |
150 |
| NCT01849952 |
Glioma |
Survival biomarker |
An observational study aiming to evaluate the correlation of miRNA-10b expression levels with patients' survival, tumor grade, and genotypic variations |
200 |
| NCT05443412 |
Prostate cancer |
Diagnostic biomarker |
An observational study assessing the possibility of artificial intelligence-assisted-based prostate cancer diagnosis based on the expression levels of miRNAs |
510 |
| NCT04965259 |
HCC |
Diagnostic biomarker |
An observational study aims to validate a panel of circulating miRNA biomarkers to develop an in-vitro diagnostic kit for the detection of early HCC |
2000 |
| NCT05431621 |
Digestive system cancers |
Diagnostic biomarker |
An observational study to establish molecular testing methods for non-invasive screening and early diagnosis of digestive system cancers through the expression levels of miRNA7 |
2430 |
| NCT05495685 |
Pancreatic cancer |
Diagnostic biomarker |
An observational study aimed at early detecting pancreatic cancer by combined assays for biomarkers of cfDNA methylation, serum protein markers, blood miRNA markers, and others |
450 |
| NCT03253107 |
GC |
Predicting biomarker of GC chemotherapy response |
An observational study to identify and validate a biomarker for the response to chemotherapy in GC |
800 |
| NCT05148572 |
HCC |
Disease recurrence biomarker, diagnostic biomarker |
An observational study aims to validate a panel of circulating miRNAs to aid in the diagnosis and prediction of recurrence in affected patients |
100 |
| NCT05556603 |
Pancreatic cancer |
Diagnostic biomarker |
An observational study aiming to evaluate the possibility of blood miRNAs for the diagnosis of pancreatic cancer patients |
7062 |
| NCT05901376 |
GC |
Diagnostic biomarker |
An observational study determining the possibility of blood miRNAs for the diagnosis of GC |
280 |
| NCT06261294 |
Lung cancer |
Diagnostic biomarker |
A two-arm, open-label, non-randomized controlled pilot study validating the circulating miRNA expression as a diagnostic biomarker |
800 |
| NCT05529251 |
Seminoma |
Predicting biomarker of seminoma treatment response |
A phase 2 study aims to validate the serum levels of miRNA-M371 association with clinical stage, primary tumor size, and response to treatment in patients. |
90 |
| NCT06060873 |
Malignant testicular germ cell tumor |
Diagnostic biomarker |
A phase 2 study of serum miRNA-371 in the diagnosis of the affected patients |
418 |
| NCT04914026 |
Testicular cancer |
Predicting biomarker of testicular cancer chemotherapy response, disease recurrence biomarker |
An observational study determining the miRNA-371 as a marker for disease activity and as a tool to monitor the impact of chemotherapy and detection of recurrence in patients |
350 |
| NCT05089747 |
Solid tumors |
Treatment efficacy biomarker, diagnostic biomarker, prognostic biomarker |
An observational study aims to analyze blood miRNAs to validate the circulating miRNAs as biomarkers for cancer diagnosis, treatment efficacy, and disease progression. |
6000 |
| NCT05417048 |
BC |
Diagnostic biomarker |
A non-randomized cohort study was carried out at a single center to assess the clinical diagnostic performance of glycosylated extracellular vesicles and their contents in the early detection of BC. |
420 |
| NCT04406831 |
Pancreatic cancer |
Treatment efficacy biomarker, diagnostic biomarker, prognostic biomarker |
An observational study aims to ascertain the utility of miRNA as a biomarker in predicting treatment response and providing prognostic information for patients. |
200 |
| NCT06206603 |
CRC |
Diagnostic biomarker |
An observational study determining blood miRNAs as a potential biomarker for the diagnosis of patients |
400 |
| NCT04906330 |
BC |
Diagnostic biomarker |
An observational study evaluating miRNAs for early BC detection |
500 |
| NCT06203496 |
Glioma |
Disease recurrence biomarker |
An observational study aims to describe plasma levels of pro-oncogenic miRNAs, after surgery for grade 4 glioma, in order to assess whether they can be utilized to detect false-positive recurrences on MRI |
60 |
| NCT05292573 |
Endometrial malignancy |
Outcome prediction biomarker |
A phase 3 study aims to evaluate miRNAs as biomarkers for predicting future endometrial malignancy in women with endometrial hyperplasia without atypia |
1000 |
| NCT06314971 |
CRC |
Diagnostic marker of disease recurrence |
An observational study aims to validate tissue miRNA correlation with tumor recurrence after curative resection. |
200 |
| NCT06154317 |
MM |
Treatment efficacy biomarker |
An observational study seeks to validate target therapy efficacy in MM cells from affected individuals by miRNAs released from B Cells |
30 |
| NCT04113122 |
Testicular cancer |
Treatment efficacy biomarker |
An observational study aims to confirm the potential of miRNA -103, miRNA -107, and miRNA -29 tissue expression as a treatment efficacy biomarker |
192 |
Abbreviations: SCC: squamous cell carcinoma, BC: breast cancer, CRC: colorectal cancer, GC: gastric cancer, HPV: human papillomavirus, HCC: hepatocellular carcinoma, PD-L1: programmed cell death ligand 1, PC: prostate cancer, MM: multiple myeloma, miRNA: microRNA, MRI: magnetic resonance imaging.
Interplay between miRNAs and tumor-associated innate immune cells
Tumor-associated Macrophages
Tumor-associated macrophages (TAMs), originating from tissue-resident macrophages or bone-marrow-derived monocytes, stand out as critical players within the TME, substantially impacting tumor cells' progression, metastasis, angiogenesis, and even side effects of therapies. Investigations have demonstrated that they exhibit anti-tumorigenic and tumoricidal properties during the initial stages of tumor development. However, as the cancer progresses to its intermediate and advanced stages, TAMs undergo a functional transition, subsequently exerting pro-tumorigenic effects that facilitate tumor growth, invasion, and metastasis. Suppressive immune cells and immunomodulatory factors present within the TME, along with metabolic alterations within cells, induce a shift in the macrophage phenotype from an anti-tumor to a pro-tumor state.42,43
Due to TAMs' capacity to modulate both innate and adaptive immune responses, coupled with their substantial presence within the TME, they exert a profound influence on the TME's status and nowadays they are regarded as promising therapeutic targets for cancer immunotherapy strategies.44 Key treatment strategies involving TAMs include depleting pro-tumor TAMs, reprogramming them and activating anti-tumor ones, blocking their recruitment to the TME, and employing novel therapies such as nanocarrier-based approaches to directly target TAMs or modulate their molecular expression profiles.45-48
The polarization of TAMs within the TME into two distinct subsets, classically activated (M1 or pro-inflammatory subtype) and alternatively activated (M2 or immunosuppressive subtype), is a highly flexible and reversible process, which shows opposing effects on tumor progression.49 The M1 macrophage phenotype (MHC II⁺, iNOS⁺, CD86⁺,) is induced by GM-CSF, IFN-γ, LPS, and TLR activation. These cells have pro- and anti-tumor functions, producing cytokines like TNF-α, IL-1β, IL-6, and IL-12, along with ROS/NOS. M1 macrophages also promote immune cell infiltration, particularly Th1 cells, supporting anti-tumor immunity.50 On the other side, the M2 phenotype (CD163+, CD206+, Arginas1hi, and vascular endothelial growth factor (VEGF)hi) is induced mainly by CSF-1, IL-4, IL-10, IL-13, and TGF-β cytokines and activation of transcriptional factors, including signal transducer and activator of transcription 6 (STAT6), suppressor of cytokine signaling 2 (SOCS2), and peroxisome proliferator-activated receptor γ (PPAR).51,52 In the context of cancer, the effects of IL-4 and IL-13, produced by Th2 cells, on the induction of M2 macrophages have explicitly been elucidated. All these factors ultimately contribute to forming an immunosuppressive TME, facilitating angiogenesis, epithelial-mesenchymal transition (EMT), tissue remodeling, tumor progression, and an unfavorable clinical prognosis. Thus, it is evident that M2 cells mainly contribute to cancer progression by both promoting the growth and development of tumor cells and suppressing immune system responses.53 M2 macrophages can be classified into M2a, M2b, M2c, and M2d, each playing distinct functions in progression of cancer. M2a cells, induced by IL-4 and IL-13, enhance tumor growth, angiogenesis, and immune suppression through factors like TGF-β and IL-10. M2b, known as regulatory macrophages, suppress immune responses and support Treg and Th2 differentiation. M2c promotes tumor invasion, while M2d secretes pro-tumoral cytokines (e.g., IL-6, VEGF, MMPs) and aids immune evasion, collectively fostering progression of tumor.53,54
Current investigations have confirmed the impact of ncRNAs, especially miRNAs, on the polarization of macrophages.55-59 Notably, the bidirectional exchange of intercellular exosomes containing miRNAs, especially between tumor cells and TAMs, has recently emerged as a critical mediator of this process.60,61
M1 polarization
Recent research highlights the capacity of specific miRNAs derived from tumor cells to induce the differentiation of M1 macrophages. In this context, hepatocellular carcinoma (HCC) has been a focal point of investigation. Li et al62 showed that miR-98 is downregulated in HCC-associated TAMs, and its overexpression promotes polarization of M2-to-M1, decreasing invasion and EMT in HCC cells. Similarly, targeted delivery of miR-99b to TAMs induced M1 polarization via the mTOR/NF-κB pathway and suppressed M2 differentiation by inhibiting mTOR/IRF4, enhancing anti-tumor immunity in the TME.63 Additionally, HCC cells with overexpressed miR-144/miR-451a cluster exhibited a distinct macrophage polarization shift in TAMs towards the M1 phenotype, which was mediated by regulating the expression of macrophage migration inhibitory factor (MIF) and hepatocyte growth factor (HGF).64
It was shown that downregulated miR-148a expression in SW480 colorectal cancer (CRC) cells has the ability to induce THP-1 cell differentiation to M2 subtype and reduce macrophage infiltration. However, induced expression of miR-148a promoted the differentiation of THP-1 cells to M1 phenotype via targeting signal regulatory protein α (SIRPα), which further induced apoptosis in SW480 cells.65 Previous research has identified miR-302a as a key player in the regulation of M1 macrophage polarization in glioma tumors. JMJD1C, a histone demethylase, affects macrophage polarization by regulating the miR-302a/N6-adenosine-methyltransferase 70kDa subunit (METTL3)/suppressor of cytokine signaling 2 (SOCS2) pathways in glioblastoma. In fact, by miR-302a overexpression, M1 polarization was induced, and glioma progression was suppressed.66 In a study on mouse breast cancer (BC) model by Moradi-Chaleshtori et al,67 it was found that 4T1 BC cell-extracted exosomes containing miR-33 have the ability to shift M2 to M1 subtype, in such a way that they got the potential to suppress 4T1 cells growth and progression. Recently, Yang et al68 revealed that pancreatic ductal adenocarcinoma (PDAC) samples had low levels of miR-506, which, upon restoration, was able to reorient M2 macrophages towards an M1 phenotype by targeting STAT3. This regulatory pathway facilitated the infiltration of CTLs in TME and enhanced the response to anti-programmed cell death protein 1 (PD-1) immunotherapy. Investigation about prostate cancer showed that exosomes containing miR-203 induce the polarization of M1 macrophages and suppress the progression of prostate cancer tumor cells69 (Table 2, Fig. 2).
Table 2.
miRNAs and M1 polarization in the context of cancer
|
miRNA
|
Model
|
Intervention/
Expression
|
TAMs
|
Outcomes
|
Ref.
|
| miR-98 |
Human |
miR-98 mimic/inhibitor transfected into TAMs |
PBMCs-derived monocytes |
Induced expression of miR-98:
|
62
|
| miR-99b |
Mouse |
miR-98 agomir/antagomir transfected to TAMs |
BM-derived
macrophages of HCC-bearing
mice |
Induced expression of miR-99:
-
Improved M1 polarization, phagocytosis, and antigen presentation via targeting mTOR/IRF4 expression
-
Increased CD8+T cells, and decreased MDSCs and Treg cells
-
Inhibited tumor growth
|
63
|
miR-144
miR-451a |
Human |
HCC and para-tumor tissues |
TAMs within tumor tissues |
Induced expression of miR-14/miR-451a:
-
Facilitated M1 polarization via targeting HGF and MIF
-
Improved CD8+T cells and reduced Tregs infiltrating
-
Increased anti-tumor cytokines and molecules
|
64
|
| Mouse |
miR-144/miR-451a transfected to HCC cells |
BM-derived
macrophages |
Induced expression of miR-14/miR-451a:
|
| miR-148a |
Human |
miR-148a mimic/inhibitor transfected to macrophages |
THP-1 cell line-derived
macrophages |
Induced expression of miR-148a:
|
65
|
| Mouse |
miR-148a mimic/inhibitor transfected to macrophages |
THP-1 cell line-derived
macrophages |
Induced expression of miR-148a:
|
| miR-302a |
Human |
JMJD1C transfected Glioma cell lines |
CD14+PBMCs |
Induced expression of miR-302-a:
|
66
|
| Mouse |
JMJD1C transfected to Glioma cell line |
CD11b+TAMs within tumor tissues |
Induced expression of miR-302-a:
|
| miR-33 |
Mouse |
BC cells-derived exosomes |
Peritoneal macrophages |
Induced expression of miR-33:
-
Converted M2 to M1 phenotype
-
Reduced proliferation, invasion, and migration of BC cells
|
67
|
| miR-506 |
Mouse |
miR-506 mimic was injected intraperitoneally |
- |
Induced expression of miR-506:
-
Reduced M2/M1 ratio
-
Reduced tumor size and weight
-
Improved survival
-
Improved CTLs and reduced Tregs infiltrating
-
Improved response to anti-PD-1 therapy
|
68
|
| miR-203 |
Human |
prostate cancer cell line-derived exosomes
miR-203 agomir/antagomir transfected to macrophages and prostate cancer cell line |
Human macrophages |
Induced expression of miR-203:
-
Suppressed prostate cancer cell proliferation, migration, and invasion
-
Induced prostate cancer cell apoptosis
|
69
|
| Mouse |
miR-203 agomir/antagomir transfected to TAMs |
prostate cancer -bearing
mice |
Induced expression of miR-203:
|
Abbreviations: miR: microRNA, TAMs: Tumor-associated macrophages, PBMCs: peripheral blood mononuclear cells, HCC: hepatocellular carcinoma, EMT: epithelial-mesenchymal transition, BM: bone marrow, mTOR: the mammalian target of rapamycin, IRF4: interferon regulatory factor 4, MDSCs: myeloid-derived suppressor cells, Tregs: regulatory T cells, HGF: hepatocyte growth factor, MIF: macrophage migration inhibitory factor, SIRPα: signal regulatory protein α, METTL3: N6-adenosine-methyltransferase 70kDa subunit, SOCS2: suppressor of cytokine signaling 2, BC: breast cancer; CTLs: cytotoxic T cells, PD-1: programmed cell death protein 1.
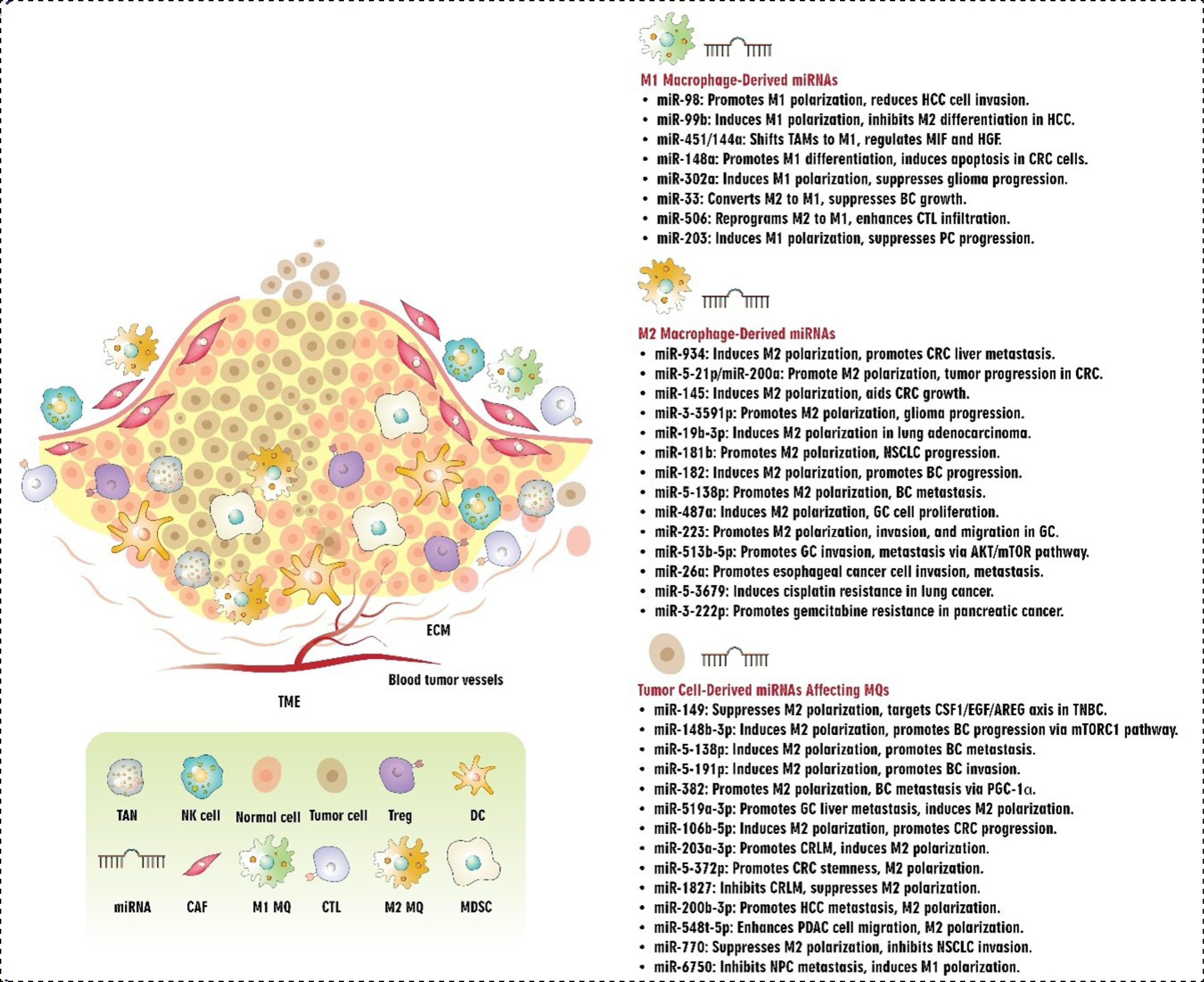
Fig. 2.
Interplay between miRNAs and Macrophages in TME. Abbreviations: TME: tumor microenvironment, ECM: extracellular matrix, miRNA: microRNA, TAN: tumor-associated neutrophils, NK cell: natural killer cell, Treg: T regulatory cell, DC: dendritic cell, CAF: cancer-associated fibroblast, MDSC: myeloid-derived suppressor cells, CTL: cytotoxic T cells, MQ: macrophage, HCC: hepatocellular carcinoma, TAM: tumor-associated macrophages, MIF: macrophage migration inhibitory factor, HGF: hepatocyte growth factor, CRC: colorectal cancer, BC: breast cancer, PC: prostate cancer, NSCLC: non-small cell lung cancer, GC: gastric cancer, TNBC: triple-negative breast cancer, PGC-1α: peroxisome proliferator-activated receptor γ coactivator-1α, CRLM: CRC liver metastasis, PDAC: pancreatic ductal adenocarcinoma, NPC: nasopharyngeal carcinoma.
.
Interplay between miRNAs and Macrophages in TME. Abbreviations: TME: tumor microenvironment, ECM: extracellular matrix, miRNA: microRNA, TAN: tumor-associated neutrophils, NK cell: natural killer cell, Treg: T regulatory cell, DC: dendritic cell, CAF: cancer-associated fibroblast, MDSC: myeloid-derived suppressor cells, CTL: cytotoxic T cells, MQ: macrophage, HCC: hepatocellular carcinoma, TAM: tumor-associated macrophages, MIF: macrophage migration inhibitory factor, HGF: hepatocyte growth factor, CRC: colorectal cancer, BC: breast cancer, PC: prostate cancer, NSCLC: non-small cell lung cancer, GC: gastric cancer, TNBC: triple-negative breast cancer, PGC-1α: peroxisome proliferator-activated receptor γ coactivator-1α, CRLM: CRC liver metastasis, PDAC: pancreatic ductal adenocarcinoma, NPC: nasopharyngeal carcinoma.
M2 polarization
In addition to miRNAs involved in M1 polarization, studies have elucidated the pivotal role of specific miRNAs in inducing the differentiation of M2 macrophages and promoting tumor progression. miR-934 present within the exosomes derived from CRC cell lines induced M2 polarization via phosphatase and tensin homolog (PTEN)/phosphoinositide 3-kinases (PI3K)/protein kinase B (PKB) signaling pathway, which in turn promoted CRC liver metastasis (CRLM) via chemokine CXCL-13/ CXCR5/NF-κB/p65/miR-934 positive feedback loop.70 Yin et al71 investigated miRNA profiles within exosomes derived from CRC cells. They found that miR-21-5p and miR-200a modulate the polarization of M1 to M2 macrophages and even their PD-L1 expression via PTEN/AKT and SOCS1/STAT1 pathways. Additionally, they noticed that this polarization to the M2 subtype made an immunosuppressive TME and promoted tumor progression. Another study regarding CRC showed that miR-145 within CRC cell-derived exosomes affected the expression of histone deacetylase 11 and induced polarization of THP-1 cells to the M2 subtype. Moreover, polarized M2 macrophages helped CRC cells grow and tumor enlargement.72
miR-3591-3p was found to be highly present in both cerebrospinal fluid (CSF) and exosomes derived from glioma cells, which promoted M2 polarization and tumor progression.73 Regarding lung adenocarcinoma, it was shown that miR-19b-3p within lung adenocarcinoma-derived exosomes induced polarization of TAMs to M2 cells via positive effects on STAT3 and LINC00273 in THP-1 cells.74 Additionally, miR-181b showed an overexpression in non-small cell lung cancer (NSCLC) patients' serum and NSCLC cells-derived exosomes, which induced M2 polarization through the JAK2/STAT3 pathway and promoted NSCLC progression.75
In a study regarding BC, Ma et al. found that miR-182 knockout inmice and macrophages hinders the development of M2-like TAMs and tumor progression. Through TGF-β secretion, BC cells induced the expression of miR-182 in TAMs, which targeted TLR4 and suppressing NF-κB shifted TAMs to the M2 subtype.76 Another study revealed that exosomes derived from MDA-MB-231 cells, enriched with miR-138-5p, altered the differentiation of THP-1 cells towards an M2-like phenotype by modulating KDM6B (Lysine Demethylase 6B) expression. This resulted in the enhanced metastatic potential of BC cells to the lung77 (Table 3).
Table 3.
miRNAs and M2 polarization in the context of cancer
|
miRNA
|
Model
|
Intervention/ Expression
|
TAMs
|
Outcomes
|
Ref.
|
| miR-934 |
Human |
CRC cell lines-derived exosomes
miR-934 mimic/inhibitor transfected to macrophages and CRC cell lines |
THP-1 cell line/BM-derived
Macrophages |
Induced expression of miR-934:
|
70
|
| Mouse |
CRC cell lines treated with condition media of THP-1 and miR-934 mimic/inhibitor transfected CRC cell lines |
THP-1 cell line-derived
macrophages |
Induced expression of miR-934:
|
| miR-21-5p miR-200a |
Human |
CRC cell lines-derived exosomes
miR-21-5p miR-200a mimic/inhibitor transfected to macrophages and CRC cell lines |
THP-1 cell line-derived
macrophages |
Induced expression of miR-21-5p miR-200a:
|
71
|
| Mouse |
Exosomes containing high miR-21-5p miR-200a expression |
Peritoneal macrophages |
Induced expression of miR-21-5p miR-200a:
|
| miR-145 |
Human |
miR-145 mimic/antagomir-transfected CRC cell lines-derived exosomes |
THP-1 or
NOMO-1 cell lines-derived
macrophages |
Induced expression of miR-145:
|
72
|
| Mouse |
Co-injection of miR-145 mimic/antagomir-transfected macrophages to CRC cell–xenografted mice |
THP-1 or
NOMO-1 cell lines-derived
macrophages |
Induced expression of miR-145:
|
| miR-3591-3p |
Human |
Glioma cell lines-derived exosomes
miR-3591-3p mimic/inhibitor transfected to macrophages |
THP-1 cell line-derived
macrophages |
Induced expression of miR-3591-3p:
-
Facilitated M2 polarization via targeting CBLB
-
Increased macrophage infiltration
-
Promoted glioma invasion and migration
|
73
|
| Mouse |
miR-3591-3p transfected to macrophages |
THP-1 cell line-derived
macrophages |
Induced expression of miR-3591-3p:
-
Increased tumor size
-
Reduced overall survival
|
| miR-19b-3p |
Human |
lung adenocarcinoma cell-derived exosomes
miR-19-3p mimic/inhibitor transfected to macrophages |
THP-1 cell line-derived
macrophages |
Induced expression of miR-19-3p:
|
74
|
| Mouse |
Coinjection of miR-19-3p mimic/inhibitor-transfected macrophages to lung adenocarcinoma cell–xenografted mice |
THP-1 cell line-derived
macrophages |
Induced expression of miR-19-3p:
|
| miR-181b |
Human |
NSCLC cell-derived exosomes |
THP-1 cell line-derived
macrophages |
Induced expression of miR-181b:
-
Facilitated M2 polarization via targeting JAK2/STAT3 axis
-
Promoted NSCLC cell proliferation, migration, and invasion.
|
75
|
| miR-182 |
Human |
miR-182 mimics transfected to macrophages |
U937 and THP-1 cell lines-derived
macrophages |
Induced expression of miR-181b:
|
76
|
| Mouse |
BC conditioned medium |
BM-derived macrophages from miR-182 KO
mice |
Induced expression of miR-181b:
|
| miR-138-5p |
Human |
BC cell lines-derived exosomes
miR-138-5p mimics transfected to macrophages |
THP-1 cell line-derived
macrophages |
Induced expression of miR-138-5b:
|
77
|
| Mouse |
BC cell lines-derived exosomes |
Raw264.7 cell line-derived
macrophages |
Induced expression of miR-138-5b:
|
Abbreviations: miRNA: microRNA, CRC: colorectal cancer, TAMs: tumor-associated macrophages, BM: bone marrow, CXCL13: C-X-C motif chemokine ligand 13, CXCR5: C-X-C motif chemokine receptor 5, NFκB: nuclear factor kappa B, PD-L1: programmed death-ligand 1, HDAC11: histone deacetylase 11, CBLB: Casitas B-lineage lymphoma-B, NSCLC: non-small cell lung cancer, JAK2: Janus kinase 2, STAT3: signal transducer and activator of transcription 3, TLR4: Toll-like receptor 4, NF-κB: nuclear factor kappa-light-chain-enhancer of activated B cells, BC: breast cancer, KDM6B: lysine demethylase 6B.
Regarding the relationship between miRNAs and the differentiation of M2a, M2b, M2c, and M2d cells in cancer, no studies have been conducted to date, and existing findings are restricted to normal physiological conditions. Furthermore, using next-generation sequencing (NGS), Jiménez et al reported that M2a-polarized macrophage has a high expression of miR-500a-5p and miR-502-3p, as well as reduced level of miR-181-5p. Additionally, they showed that the M2c subtype has a high level of miR-21-5p, miR-22-3p, and miR-146b-5p, while miR-200a-3p and miR-339-3p expression were decreased.78 Additionally, there is evidence linking miR-222 to the differentiation of M2b macrophages.79 Collectively, these findings suggest a clear relationship between the regulatory effects of miRNAs and the polarization of M2 subtypes. However, these studies are predominantly conducted under normal physiological conditions. Further research is warranted to investigate how miRNAs influence M2 subtype polarization within the TME, where factors such as hypoxia, metabolic changes, and cytokine gradients are likely to modulate their effects (Fig. 2).
Tumor growth and metastasis
As previously mentioned, miRNAs have been found to play a crucial role in shaping the function of TAMs in the TME, allowing them to acquire novel phenotypes that either promote or suppress tumor cell growth, progression, migration, and invasion. Based on the available findings and evidence, altered expression of miRNAs, whether inside macrophages (indirectly) or through exosome delivery to tumor cells (directly), could affect tumor cell progression or suppression. In the following sections, the miRNAs derived from tumor cells and macrophages and the effects they have on each other are discussed.
a. Tumor-derived miRNAs
It was shown that miR-149 was downregulated in triple-negative breast cancer (TNBC) cells, and its altered expression has a relation with macrophage infiltration and tumor progression. miR-149 overexpression in TNBC cells suppressed BC cells' communication with THP-1 cells and impaired their polarization to M2 phenotype by targeting the CSF1/EGF/AREG axis.80 Additionally, miR-148b-3p was overexpressed in exosomes produced by BC cells, which led to the reprogramming of TAMs to M2 macrophages and thereby promoted BC progression via tuberous sclerosis complex2 (TSC2) targeting and induction of the mTORC1 pathway.81 As mentioned before, miR-138-5p present in BC cell-derived exosomes induced M2 polarization and BC cell metastasis.77 Furthermore, overexpression of miR-191-5p in BC cells and their secreted exosomes and its delivery to macrophages affected their polarization to the M2 subtype via targeting the SOCS3 expression, promoting BC cell invasion and migration.82 Moreover, 4T1 BC cells-derived exosomes containing miR-33 affected the shift of M2 to M1, suppressing BC growth and metastasis.67 Another study regarding BC revealed the inhibitory effect of BC cells on miR-382 expression in TAMs, where peroxisome proliferator-activated receptor γ coactivator-1α (PGC-1α), as its target, showed increased expression and induced M2 polarization. Afterward, these M2 cells promoted BC cell metastasis and EMT.83
Regarding gastrointestinal cancer, gastric cancer (GC)-derived exosomes overexpressing miR-519a-3p displayed a vital role in promoting angiogenesis, leading to GC liver metastasis (GC-LM) via affecting intrahepatic macrophages. It was shown that delivering highly expressed miR-519a-3p within these exosomes targeted the DUSP2-MAPK/ERK pathway in macrophages, inducing their polarization to the M2 subtype and production of angiogenic factors in the liver.84 Moreover, GC-derived exosomes enriched in let-7g-5p were shown to mediate macrophage M2 polarization that is linked to tumor-promoting immunosuppression and tumor progression and SERPINE1 was identified as the main mediator responsible for let-7g-5p loading into exosomes. When taken up by macrophages, let-7g-5p stimulates a metabolic and functional transition to the M2 phenotype which enhances the pro-tumorigenic milieu. Moreover, this study defines a new mechanism for tumor cells to utilize exosomal microRNAs to reprogram immune cells, which have the possibility of being future therapeutic targets, to inhibit tumor-promoting immune modulation.85 Transferring of miR-106b-5p within exosomes derived from EMT-CRC cells induced M2 polarization via affecting PI3K/AKT/mTOR pathway and finally decreasing programmed cell death protein 4 (PDCD4) expression within macrophages, which in turn helped induction of immunosuppressive TME and promotion of tumor progression.86 miR-203a-3p showed an upregulation within CRC plasma samples and cell line exosomes, where its delivery to TAMs affected their polarization to the M2 subtype through PTEN regulation and PI3K/Akt signaling pathway activation, leading to colorectal liver metastasis.87 miR-372-5p was found to be highly expressed in exosomes released by CRC and influenced macrophage polarization towards the M2 subtype via modulating the PTEN/AKT signaling pathway. Subsequently, CXCL12 chemokine production and secretion by these M2 macrophages promoted the stemness, metastasis, and EMT of CRC cells.88 Reports indicate that the CXCL12/CXCR4 axis plays a role in the pathogenesis of various human disorders, including infectious diseases, autoimmune diseases, and cancer.89-94 Exosomes derived from umbilical cord mesenchymal stem cells enriched with miR-1827 showed a prominent relation with hindering the colorectal liver metastasis process and suppressing M2 polarization via targeting succinate receptor 1 (SUCNR1), in which this effect, in turn, inhibited CRC progression.95 One study demonstrated that miR-200b-3p, present in exosomes derived from HCC, influenced the conversion of M0 macrophages to M2 by regulating the production of IL-4 cytokines and activating the JAK/STAT signaling pathway, ultimately contributing to enhanced HCC metastasis.96 It was shown that miR-548t-5p within PDAC cells affected IL-33 expression, which improved macrophage infiltration and M2 polarization. In the following, induced M2 cells improved PDAC cell migration and invasion.97 Overexpression of miR-770 within NSCLC-derived exosomes suppressed the polarization of M2 macrophages through modulating mitogen-activated protein kinase 1 (MAP3K1) and thereby barricading NSCLC cells invasion, migration, and EMT in vitro and in vivo.98 Zhang et al. studied nasopharyngeal carcinoma-derived exosomes' miRNA profile and found that tumor-suppressor miR-6750 has an inhibitory effect on nasopharyngeal carcinoma metastasis and angiogenesis through inducing M1 macrophages and targeting mannose 6-phosphate receptor (M6PR)/ERK/MAPK/P38MAPK axis. Macrophages, as mediator cells, also showed a crucial role in delivering these exosomes to human umbilical vein endothelial cells (HUVECs) and then modulating pre-metastatic niche formation.99
b. Macrophage-derived miRNAs
Investigating macrophages involved in the progression of PDAC revealed the crucial role of TAMs-derived exosomal miR-202-5p and miR-142-5p, affecting PTEN and thereby promoting tumor cell invasion and metastasis.100 Moreover, TAM-derived exosomes containing miR-501-3p affected PDAC cell formation, invasion, and metastasis via TGF-β signaling pathway activation.101 Regarding renal cell carcinoma (RCC), it was shown that increased expression of hypoxia-inducible factor 1-alpha (HIF-1α) led to the upregulation of miR-193a-5p in macrophages, which in turn triggered the release of exosomes containing this miRNA. These exosomes were then taken up by RCC cells, where they influenced the expression of tissue inhibitor of metalloproteinases 2 (TIMP2), thereby contributing to the progression of the cancer cells.102 In another study, overexpression of miR-342-3p was found both in RCC cells and M2-derived exosomes, in which transferring of M2-derived miR-342-3p to RCC cells promoted tumor cell progression through regulation of NEDD4L (The E3 ubiquitin ligase), PI3K/AKT/mTOR signaling pathway, and finally Centrosomal protein of 55 kDa (CEP55) expression, respectively.103 Concerning gastrointestinal cancer, it was shown that M2 macrophages in the TME promoted GC cell proliferation and progression via miR-487a present in GC-derived exosomes, regulating TIA-1 (T-cell intracellular antigen 1) gene expression. Additionally, miR-223 overexpression within macrophage-derived exosomes and its transfer to GC cells induced tumor cell invasion, migration, and EMT through the regulation of the PTEN-PI3K/AKT pathway.104 Investigation regarding the anti-tumoral effect of traditional Chinese medicine called Jianpi Yangzheng Xiaozheng decoction (JPYZXZ) ended up revealing the tumor-promoting effect of miR-513b-5p present in TAMs-derived exosomes. Zhang et al105 showed that transferring miR-513b-5p within exosomes to GC cells activated the AKT/mTOR signaling pathway via PTEN suppression and ultimately induced GC cell invasion and metastasis. Mi et al106 revealed the regulatory effect of lncRNA AFAP1-AS1 on miR-26a expression in esophageal cancer. They revealed that transferring M2-derived exosomes with increased levels of lncRNA AFAP1-AS1, and decreased levels of miR-26a contributed to enhanced esophageal cancer cell invasiveness, migration, and lung metastasis. Regarding lung cancer and miRNAs present in exosomes derived from M1 macrophages, recently Peng et al107 reported that transferring of miR-let-7b-5p within M1-derived exosomes showed tumor-suppressing effects on lung cancer cells through its regulatory effects on G protein subunit gamma 5 (GNG5).
Therapy resistance
Studies conducted in recent years on chemotherapy drug resistance indicate that TAM-derived miRNAs influence the development of resistance in tumor cells. M2 polarization and co-culturing their exosomes with GC cells revealed that miR-21 plays a crucial role in the development of cisplatin resistance by regulating the PTEN/PI3K/AKT signaling pathway.108 A confirmatory study on ovarian cancer demonstrated that the up-regulation of miR-21 was not only involved in M2 polarization but also contributed to the induction of cisplatin chemoresistance through miR-21 delivery within exosomes derived from M2 macrophages.109 Hypoxic conditions in the TME of ovarian cancer were shown to increase macrophage infiltration and TAM polarization that secreted exosomes enriched with miR-223. The transfer of these exosomes from TAMs to ovarian cancer cells promoted resistance to cisplatin through the PTEN-PI3K/AKT pathway.110 Another study demonstrated that exosomal miRNA-223 derived from M2 macrophages can enhance doxorubicin resistance in GC cells by modulating the expression of the F-box and WD repeat domain-containing 7 (FBXW7) protein.111 miR-3679-5 is the other exosomal miRNA derived from M2 macrophages, in which transferring miR-3679-5 to lung cancer cells induced metabolic changes and developed resistance to cisplatin via the regulatory effect on the NEDD4L/c-Myc axis 112. Furthermore, induction of mouse M2 macrophages and investigation of their related exosomes revealed that up-regulation of miR-222-3p promoted chemoresistance to gemcitabine in pancreatic cancer cells through targeting TSC1 expression and PI3K/AKT/mTOR pathway activation113 (Figs. 2 and 3).
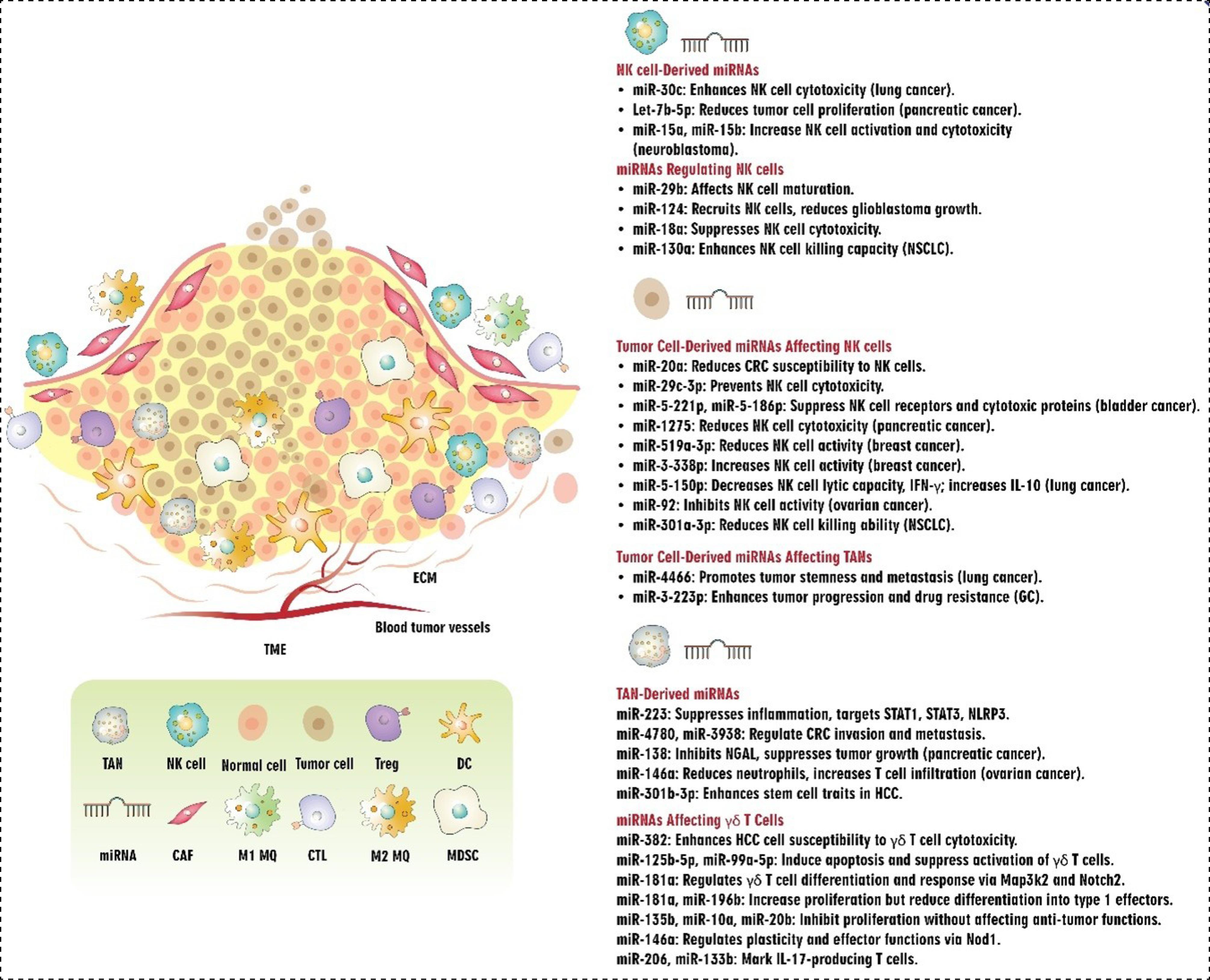
Fig. 3.
Interplay between tumor cells and macrophages in the TME mediated by miRNAs. The schematic illustrates the bidirectional interaction between tumor cells and macrophages via EVs carrying miRNAs in the TME. miRNAs regulate key signaling pathways, such as PI3K/AKT/mTOR, MAPK/ERK, JAK-STAT, and PTEN, modulating macrophage polarization into pro-inflammatory M1 or anti-inflammatory M2 phenotypes. M2 macrophages promote tumor progression, including proliferation, EMT and metastasis, angiogenesis, cancer stemness, and chemoresistance. Additionally, TAMs release EV-derived miRNAs, further enhancing tumor cell malignancy and therapy resistance. Abbreviations: EV: extracellular vesicle, EMT: epithelial-to-mesenchymal transition, TAM: tumor-associated macrophage, PI3K: phosphoinositide 3-kinases, mTOR: mammalian target of rapamycin, PTEN: phosphatase and tensin homolog, SOCS: suppressor of cytokine signaling, PGC-1α: peroxisome proliferator-activated receptor γ coactivator-1α, TGF: Transforming growth factor, TIMP2: tissue inhibitor of metalloproteinases 2, M6PR: mannose 6-phosphate receptor, VEGF: vascular endothelial growth factor, FBXW7: F-box and WD repeat domain-containing 7.
.
Interplay between tumor cells and macrophages in the TME mediated by miRNAs. The schematic illustrates the bidirectional interaction between tumor cells and macrophages via EVs carrying miRNAs in the TME. miRNAs regulate key signaling pathways, such as PI3K/AKT/mTOR, MAPK/ERK, JAK-STAT, and PTEN, modulating macrophage polarization into pro-inflammatory M1 or anti-inflammatory M2 phenotypes. M2 macrophages promote tumor progression, including proliferation, EMT and metastasis, angiogenesis, cancer stemness, and chemoresistance. Additionally, TAMs release EV-derived miRNAs, further enhancing tumor cell malignancy and therapy resistance. Abbreviations: EV: extracellular vesicle, EMT: epithelial-to-mesenchymal transition, TAM: tumor-associated macrophage, PI3K: phosphoinositide 3-kinases, mTOR: mammalian target of rapamycin, PTEN: phosphatase and tensin homolog, SOCS: suppressor of cytokine signaling, PGC-1α: peroxisome proliferator-activated receptor γ coactivator-1α, TGF: Transforming growth factor, TIMP2: tissue inhibitor of metalloproteinases 2, M6PR: mannose 6-phosphate receptor, VEGF: vascular endothelial growth factor, FBXW7: F-box and WD repeat domain-containing 7.
MDSCs
Multiple studies have highlighted the significance of miRNAs in tumor progression and metastasis, where they exert their influence by regulating the development, maturation, and functional activity of MDSCs. These miRNAs can be derived from MDSCs and also produced by tumor cells and transferred to MDSCs via extracellular vesicles (EVs) and exosomes to carry out their function. These regulatory effects eventually influence tumor growth, metastasis, and angiogenesis.
miRNAs in cancer cells primarily utilize EVs and exosomes to modulate tumor growth and manipulate the behavior of MDSCs, including their production, aggregation, and activity. The solid TME is often characterized by hypoxic conditions, which are believed to impact the biogenesis and release of tumor-derived exosomes. Guo et al found that glioma cells affect MDSCs through exosomes. Hypoxia-induced expression of miR-10a and miR-21 in exosomes derived from glioma activate and differentiate MDSCs by targeting retinoic acid-related orphan receptor α (RORα)/IκBα/NF-κB and PTEN/PI3K/AKT, respectively.114 This group's other study discovered that low oxygen levels increased the expression of miR-29a and miR-92a, which activated the proliferation and function of MDSCs by targeting high-mobility group box transcription factor 1 (Hbp1) and protein kinase cAMP-dependent type I regulatory subunit alpha (Prkar1a), respectively.115 Hypoxia also raised miR-1246 levels in glioma-derived exosomes through stimulating miR-1246 transcription and selective packaging by overexpression of POU class 5 homeobox 1 (POU5F1) and heterogeneous nuclear ribonucleoprotein A1 (hnRNPA1). Exosomal miR-1246 regulates MDSC differentiation and activation via DUSP3/ ERK-dependent manner. In glioma patients, elevated exosomal miR-1246 expression was strongly associated with poor survival and increased tumor recurrence.116 Li et al. demonstrated that a low-oxygen environment triggered oral squamous cell carcinoma cells to release exosomes containing miR-21, which in turn amplified the immune-suppressing effects of MDSCs by modulating the miR-21/PTEN/PD-L1 pathway.117 SOCS and protein inhibitor of activated STAT (PIAS) family members operate as key regulators in the JAK/STAT signaling pathway, acting to counterbalance its activity by forming a negative feedback loop.118 Jiang et al. discovered that exosomes released by tumors contained miRNAs miR-9 and miR-181a, which interacted with SOCS3 and PIAS3 to activate the JAK/STAT signaling pathway. This led to the development and expansion of early-stage MDSCs, promoting their accumulation. Moreover, when these miRNAs are transmitted to MDSCs, they can suppress T-cell expansion, trigger T-cell apoptosis, and enhance tumor growth by facilitating MDSC infiltration in vivo.119 Bruns et al reported that chronic lymphocytic leukemia (CLL) cell-derived exosomes containing miR-155 promote MDSC activation and accumulation, which suppresses T-cell activation and induces suppressive Treg; however, vitamin D therapy inhibits this process.120 miRNAs, particularly miR-107, contained within exosomes secreted by immune cells can be internalized by MDSCs. This leads to a decrease in the expression of DICER1 and PTEN genes within MDSCs, resulting in enhanced cell proliferation and activation, as well as increased production of Arginase-1, a key enzyme involved in immunosuppressive activity.121 In the context of pancreatic cancer, Basso et al. reported that PDAC-secreted exosomes containing miR-494-3p and miR-1260 mediated the suppressive function of MDSCs by boosting intracellular calcium fluxes in a Smad4-dependent way.122
Exosomes produced by MDSCs have been proven in studies to transport miRNAs and affect tumor metastasis. miR-143-3p in Granulocytic-MDSC (G-MDSC)-derived exosomes increased lung cancer cell proliferation by suppressing the integral membrane protein 2B (ITM2B) transcription and stimulating the PI3K/Akt signaling pathway.123
Research has demonstrated that miRNA in MDSCs can influence their behavior and developmental trajectory. CCAAT/enhancer binding protein (C/EBP), which includes transcription factors plays a crucial role in regulating cell cycle progression and cellular differentiation in various cell types. C/EBPα is one of the family members that controls the balance of cell proliferation and differentiation. Activation of C/EBPα promotes differentiation while inhibiting proliferation of target cells.124 miR-486 targets C/EBPα, and their expression is inversely linked. In lung cancer, miR-486 was found to be highly expressed in tumor-induced monocytic MDSCs (TM-MDSCs), and elevated miR-486 levels enhanced cell proliferation and inhibited apoptosis of TM-MDSCs.125 miR-6991-3p reduces the MDSCs proliferation and activation in the TME. miR66991-3p directly targets galectin-9, a recently discovered immunological checkpoint and activator of STAT3. Hence, miR-6991-3p has been found to act as a negative regulator of STAT3 activation.126,127 Runt-related transcription factor 1 (RUNX1) a key member of the RUNX family of transcription factors, plays a crucial role in regulating the development and function of MDSC.128 Tian et al. discovered that miR-9 reduces RUNX1 expression by targeting its 3'UTR. Overexpression of miR-9 suppresses MDSC differentiation into a mature myeloid cell, increases their immunosuppressive activity, and hence promotes tumor formation.129 miR-30a is known to activate the JAK/STAT pathway by targeting SOCS3, promoting the formation of MDSCs in B-cell lymphoma. Xu et al discovered that miR-30a, which is highly expressed in MDSCs from mice with B-cell lymphoma, directly targets the 3′UTR of the SOCS3 gene. This leads to increased levels of Arginase-1, IL-10, and ROS, hence promoting MDSC differentiation, infiltration, suppressive capabilities, and tumor advancement.130 Some miRNAs exhibit dual regulatory effects. miR-155, for example, has dual regulatory effects and can operate as both an oncogene and a tumor suppressor depending on the cellular environment and target genes. SOCS1, one of the members of the SOCS family, has an important role in the negative regulation of the JAK/STAT pathway.131 Chen et al found that miR-155 regulates MDSCs by directly targeting SOCS1, which eliminates SOCS1-mediated regulation on the JAK/STAT pathway, leading to MDSC accumulation and immunosuppressive function.132 In line with these findings, Li et al. found that in an animal model of lung cancer, the co-expression of miR-155 and miR-21 enhances the proliferation and immunosuppressive capacity of MDSCs by simultaneously targeting SHIP-1 and PTEN, ultimately resulting in excessive STAT3 activation.133 On the contrary, Wang et al. provided evidence that MDSCs that lack miR-155 exhibit enhanced immunosuppressive function and are more effective at facilitating the growth of solid tumors. Their results revealed that HIF-1α, which is directly affected by miR-155, was upregulated in MDSCs with miR-155 deficiency, increasing the expression of chemokine ligands and accelerating MDSC migration into the tumor.134
Tumor-derived cytokines and inflammatory factors are thought to impact MDSC. Tumor-derived GM-CSF stimulates the expression of miR-200c in MDSCs, which are recruited to the TME. miR-200c enhances MDSC proliferation and immunosuppressive function by inhibiting FOG2 and PTEN, thereby activating the PI3K/Akt pathway.135 Liu et al observed that tumoral cell-derived TGF-β1 upregulates miR-494 in tumor-related MDSCs. miR-494 decreases PTEN expression, which enhances MDSC infiltration into the tumor site mediated via CXCR4. Moreover, PTEN downregulation stimulates the PI3K/Akt pathway, which changes the intrinsic apoptotic/survival signal, thus contributing to the accumulation of MDSCs in tumor tissues136 (Table 4, Fig. 4).
Table 4.
miRNA/MDSC interplay in the context of cancer
|
miRNA
|
Model
|
Intervention/ Expression
|
MDSCs
|
Outcomes
|
Ref.
|
| miR-10a and miR-21 |
Mouse |
Glioma cell lines derived exosomes |
Mouse MDSCs |
Hypoxic conditions enhance the expansion and immunosuppressive function of MDSCs through the miR-10a/ RORα/IκBα/NF-κB and miR-21/ PTEN/PI3K/AKT pathways |
114
|
| miR-29a and miR-92a |
Mouse |
Glioma cell lines derived exosomes |
Mouse MDSCs |
Hypoxic conditions upregulate the expression of miR-29a and miR-92a, which enhance the proliferation and immunosuppressive activity of MDSCs by targeting Hbp1 and Prkar1a |
115
|
| miR-1246 |
Human |
Glioma patients |
PBMC-derived MDSCs |
Hypoxic conditions increase miR-1246 levels in glioma-derived exosomes, which in turn drives the differentiation and activation of MDSCs |
116
|
| miR-21 |
Mouse |
miR-21 in SCC-VII (mouse squamous cell carcinoma cell line)-derived exosomes |
Mouse splenic MDSCs |
Hypoxic conditions enhance the suppressive effect of MDSCs on γδ T cells through a miR-21/PTEN/PD-L1 axis. |
117
|
| Human |
miR-21 in Cal-27 cells-derived exosomes |
PBMC-derived MDSCs |
| miR-9 and miR-181a |
Mouse |
miR-9 and miR-181a mimics or inhibitors transfected into eMDSCs |
Mouse eMDSCs (CD11b+Gr1−) |
Promote eMDSCs expansion and development by activation of the JAK/STAT signaling pathway via inhibiting SOCS3 and PIAS3 |
119
|
| Human |
miR-9 and miR-181a mimics or inhibitors transfected into eMDSCs |
Human eMDSCs |
| miR-155 |
CLL patients |
miR-155 in CLL cell-derived exosomes |
PBMC-derived MDSCs |
promote MDSC activation and accumulation |
120
|
| miR-107 |
Human |
Gastric cancer and gastric cell lines-derived exosomes |
PBMC-derived MDSCs |
Induce the expansion and activation of MDSCs by targeting DICER1 and PTEN |
121
|
| miR-494-3p and miR-1260 |
pancreatic cancer cell lines |
miR-494-3p and miR-1260 in PDAC-derived exosomes |
PBMC-derived MDSCs |
Enhance expansion and immunosuppressive function of MDSCs by boosting intracellular calcium fluxes in a Smad4-dependent manner |
122
|
| miR-143-3p |
Lung cancer |
G-MDSC-derived exosomes |
G-MDSCs |
Increase proliferation of lung cancer cells by targeting ITM2B |
123
|
| miR-486 |
Mouse |
miR-486 in tumor-induced M-MDSCs |
Mouse CD11b+Gr1+Ly6G−Ly6Chi/ +hi/ +MDSCs (M-MDSCs) |
Promote proliferation and inhibit apoptosis of M-MDSCs by targeting C/EBPα |
125
|
| miR-6991-3p |
Mouse |
miR-6991-3p mimic and antagomir transfected into MDSCs |
Mouse-derived MDSCs |
Suppress the expansion and activation of MDSCs |
126
|
| miR-9 |
Mouse |
miR-9 mimics or antagomirs transfected into MDSCs isolated from spleens of tumor-bearing mice |
Mouse MDSCs |
Inhibit the differentiation and enhance immunosuppressive activity of MDSCs by targeting RUNX1 |
129
|
| miR-30a |
Mouse |
miR-30a mimics transfected into bone marrow cells of mice |
Mouse MDSCs |
Promote differentiation, infiltration, and immunosuppressive function of MDSCs by targeting SOCS3 |
130
|
| miR-155 |
Mouse |
miR-155 knockout mice |
Mouse MDSCs |
Enhance the accumulation of functional MDSCs in the TME by targeting SOCS1 |
132
|
| miR-155 and miR-21 |
Mouse |
miR-155 and miR-21 mimics or inhibitors transfected into bone marrow cells of mice |
Mouse MDSCs |
Boost expansion and immunosuppressive activity of MDSCs by targeting SHIP-1 and PTEN |
133
|
| miR-155 |
Mouse |
miR-155 knockout mice |
Mouse MDSCs |
miR-155 deficiency enhances the recruitment and immunosuppressive functions of MDSCs in TME |
134
|
| miR-200c |
Mouse |
miR-200c in tumor-associated MDSCs |
Mouse MDSCs |
GM-CSF induces miR-200c in tumor-associated MDSCs, which in turn promote the expansion and immune suppressive activity of MDSCs via targeting PTEN and FOG2 |
135
|
| miR-494 |
Mouse |
miR-494 in tumor-associated MDSCs |
Mouse MDSCs |
TGF-β1 upregulates the expression of miR-494, which in turn induces the accumulation and activity of MDSCs by targeting PTEN and activating the PI3K/Akt pathway |
136
|
Abbreviations; MDSC: Myeloid-Derived Suppressor Cell, eMDSC: early-stage Myeloid-Derived Suppressor Cell, RORα: Retinoic acid-related Orphan Receptor α, IκBα: NF-Kappa-B Inhibitor Alpha, NF-κB: Nuclear Factor Kappa B, PTEN: Phosphatase and Tensin homolog, PI3K: Phosphoinositide 3-kinase, Hbp1: High-mobility group box transcription factor 1, Prkar1a: Protein kinase cAMP-dependent type I regulatory subunit alpha, PBMC: Peripheral Blood Mononuclear Cell, PD-L1: Programmed Death-Ligand 1, JAK: Janus Kinase, STAT: Signal Transducer and Activator of Transcription, SOCS: Suppressor of Cytokine Signaling, TME: tumor microenvironment, PIAS3: Protein Inhibitor of Activated STAT 3, CLL: Chronic Lymphocytic Leukemia, PDAC: Pancreatic Ductal Adenocarcinoma, Smad4: Smad Family Member 4, ITM2B: Integral Membrane Protein 2B, C/EBPα: CCAAT/Enhancer Binding Protein α, RUNX1: Runt-related transcription factor 1, SHIP-1: SH2-containing Inositol-5'-Phosphatase 1, FOG2: Friend Of Gata 2, TGF-β1: Transforming Growth Factor β1.
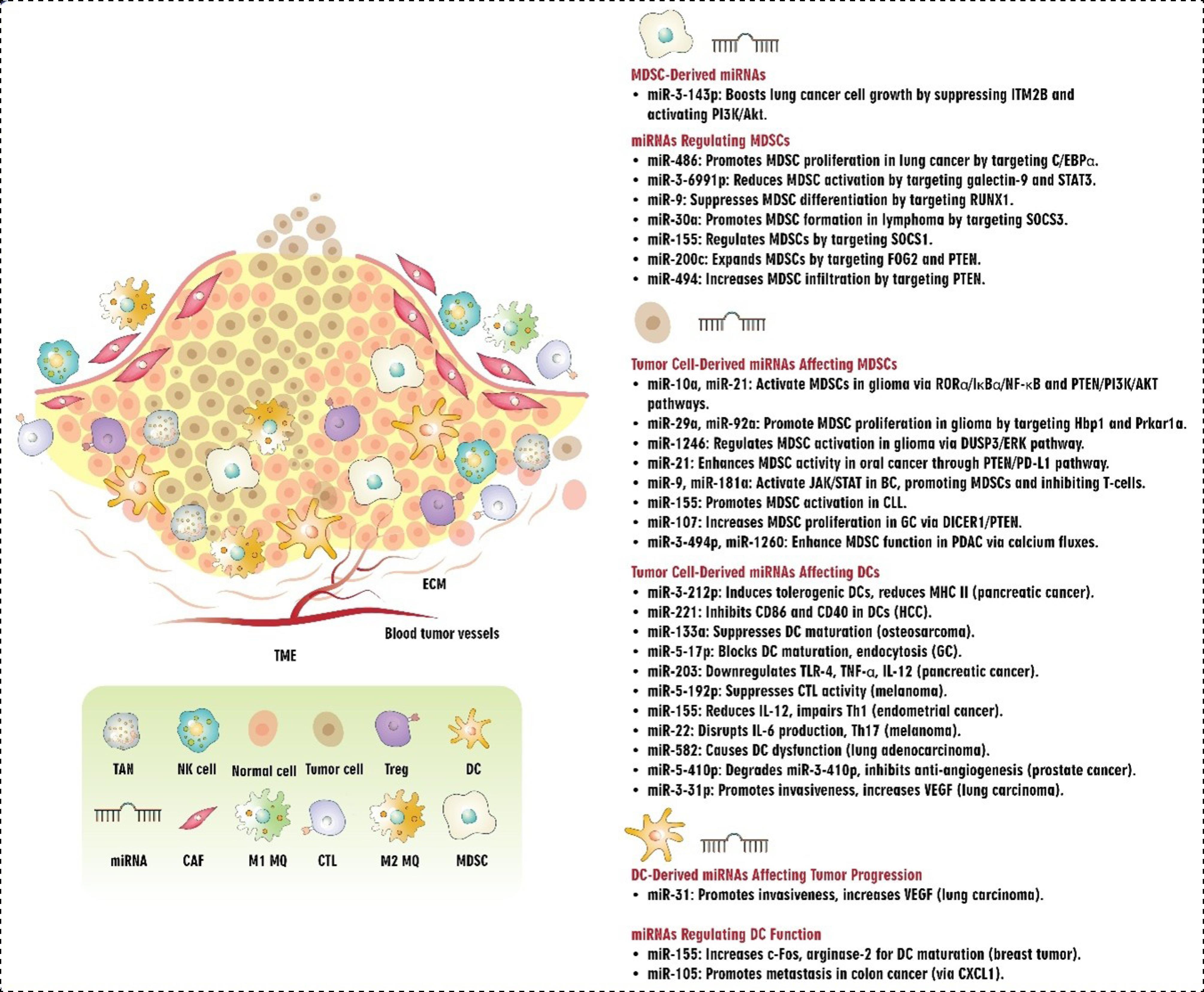
Fig. 4.
Interplay between miRNAs and MDSCs/DCs in TME.Abbreviations: TME: tumor microenvironment, ECM: extracellular matrix, miRNA: microRNA, TAN: tumor-associated neutrophils, NK cell: natural killer cell, Treg: T regulatory cell, DC: dendritic cell, CAF: cancer-associated fibroblast, MDSC: myeloid-derived suppressor cells, CTL: cytotoxic T cells, MQ: macrophage, HCC: hepatocellular carcinoma, STAT: signal transducer and activator of transcription, RUNX1: runt-related transcription factor 1, SOCS: suppressor of cytokine signaling, PTEN: phosphatase and tensin homolog, PD-L1: programmed cell death ligand 1, GC: gastric cancer, CLL: chronic lymphocytic leukemia, MHC: major histocompatibility complex, PDAC: pancreatic ductal adenocarcinoma, TLR: Toll-like receptor, TNF: tumor necrosis factor, VEGF: vascular endothelial growth factor.
.
Interplay between miRNAs and MDSCs/DCs in TME.Abbreviations: TME: tumor microenvironment, ECM: extracellular matrix, miRNA: microRNA, TAN: tumor-associated neutrophils, NK cell: natural killer cell, Treg: T regulatory cell, DC: dendritic cell, CAF: cancer-associated fibroblast, MDSC: myeloid-derived suppressor cells, CTL: cytotoxic T cells, MQ: macrophage, HCC: hepatocellular carcinoma, STAT: signal transducer and activator of transcription, RUNX1: runt-related transcription factor 1, SOCS: suppressor of cytokine signaling, PTEN: phosphatase and tensin homolog, PD-L1: programmed cell death ligand 1, GC: gastric cancer, CLL: chronic lymphocytic leukemia, MHC: major histocompatibility complex, PDAC: pancreatic ductal adenocarcinoma, TLR: Toll-like receptor, TNF: tumor necrosis factor, VEGF: vascular endothelial growth factor.
DCs
In the context of antigen presentation, DCs are regarded as the most professional antigen-presenting cells within the human body.137 DCs play a pivotal role in linking the innate and adaptive immune systems, efficiently activating naive T cells, and upholding the central aspect of anti-tumor immunity.137 Nevertheless, the appropriate function of DCs is compromised by tumor-related miRNAs.138 The transfer of miR-212-3p from pancreatic cancer-derived exosomes to DCs leads to reduced expression of RFXAP. This decrease in RFXAP results in a downregulation of MHC II expression, ultimately inducing a tolerogenic DC phenotype.139 It has been elucidated that miR-221 suppresses the expression of CD86 and CD40 on DCs that are co-cultured with HCC cells by regulating IP10.140 The maturation and activation of spleen DCs were suppressed by the miR-133a mimic in an osteosarcoma mouse model via regulating the Notch-RBP-J signaling pathway, while conversely, the miR-133a inhibitor was found to stimulate these processes.141 Studies have highlighted that the downregulation of c-Fos and Arginase-2, both identified as targets of miR-155, is critical for the maturation and functional capabilities of DCs.142,143 Accordingly, the expression levels of c-Fos and arginase-2 are increased in lymph node DCs of miR-155−/− breast tumor-bearing mice, underscoring the critical role of miR-155 expression for the efficient maturation process of DCs in breast cancer.144 The internalization of GC-derived miR-17-5p by immature DCs can impede the expression of maturation markers, including CD80, CD86, and MHC-II, and endocytosis activity of DCs stimulated by lipopolysaccharide, hence supporting GC progression.145 Exosomes released by pancreatic cancer cells containing miR-203 can reduce the expression of TLR-4, activation of NF-kB signaling pathway, and production of downstream cytokines like TNF-α and IL-12 in DCs.146 Under hypoxic conditions, melanoma cells can release miR-192-5p into the extracellular space through a mechanism involving Connexin-43 (Cx43)-mediated gap junctions, which DCs and CTLs can then take up. The transfer of miR-192-5p to CTLs through this mechanism leads to the suppression of CTL-mediated cytotoxic activity via downregulating ZEB2, a transcription factor involved in the expression of granzyme A.147 According to Jia et al, the translation of the p38 (a vital member of the MAPK14 family proteins) gene is impaired by miR-155, leading to a decrease in the ability of DCs to secrete IL-12 and polarize Th1 cells. Consequently, this process diminishes the function of DCs in inducing anti-tumor immunity in an endometrial cancer mouse model.148 In a melanoma mice model, miR-22 has been discovered to downregulate the p38 gene post-transcriptionally by inhibiting mRNA translation. This downregulation subsequently disrupted the production of DC-derived IL-6 and the stimulation of Th17 cells.149 Guo et al have provided evidence suggesting that miR-582 and its target CD1B may have significant implications in the dysfunction of DCs and could potentially be associated with clinical outcomes in advanced lung adenocarcinoma patients.150 All of these alterations in DCs can lead to the escape of the tumor cells from the immune surveillance. On the other hand, tumor-associated miRNAs have the ability to manipulate DCs and exploit their plasticity in order to facilitate the progression of tumors. In line with this, Hsu et al. have found that the CXCL1, which is highly prevalent in DCs derived from colon cancer patients, as well as SW620-conditioned tumor-associated DCs, can promote cancer stem cell characteristics.151
It is worth noting that CXCL1 enhances the metastatic capability of colon cancer cells by promoting cell migration, upregulating matrix metalloproteinase-7 expression, and inducing EMT via enhancing miR-105 in colon cancer cells through a paracrine mechanism.151 The presence of CXCL1 is associated with an increase in the expression of potential oncogenes in colon cancer, specifically PTHLH, TYRP1, FOXO1, TCF4, and ZNF880.151 The prostate cancer cell antigens induced the DCs to generate miR-410-3p, which is a highly complementary counterpart of PC-related miR-410-5p.152 miR-410-5p can enter into the DCs and this internalized miR-410-5p caused the degradation of miR-410-3p via base pairing mechanism by argonaute-2, leading to the inhibition of its function in suppressing tumor angiogenesis.152 Pyfferoen and colleagues have provided evidence indicating that the presence of hypoxia stimulates the expression of miR-31 in myeloid DCs.153 Accordingly, the upregulation of miR-31-3p in DCs results in the alteration of cellular morphology in lung carcinoma cells, leading to a decrease in sphericity and the emergence of filopodia-like protrusions.153 These changes in shape are characteristic of invasive tendencies.153 Besides, both miR-31-3p overexpression and exposure to hypoxia were shown to elevate the secretion of VEGF by DCs153 (Table S1, Fig. 4).
NK cells
NK cells, which are the innate immune system's first line of defense, use their cytotoxic and immune-regulatory abilities to combat tumors. The observed effects result from the binding of specific molecules produced by cancerous or stressed cells to receptors on the surface of those cells.154 NKG2D and its ligands, two key activators, UL16 Binding Proteins 1–6 (ULBP1–6) and MHC class I chain-related proteins A and B (MICA/B), modulate the cytotoxic potential of NK cells against cancer cells.155 While the overexpression of NKG2D ligands in cancer cells enhances the antitumor response mediated by NKG2D, reducing or eliminating NKG2D in mice impairs their ability to recognize and attack tumor cells.156,157
miRNAs play a role in regulating the capacity of NK cells to eradicate cancer cells. In this regard, it has been demonstrated that miR-20a plays a regulatory role in determining the sensitivity of CRC cells to NK cell-mediated attack by targeting MICA.158 Also, it was reported that pterostilbene-induced reduction of miR-20a in prostate cancer cells may raise MICA/B expression and decrease TGF-β1 production, which in turn may improve NK cell-mediated cytotoxicity against prostate cancer cells and provide a viable strategy for boosting anti-PC-immune-responses.159 Studies have demonstrated that miR-29b, abundantly expressed in NK cells, specifically regulates their function by inhibiting the activity of the transcription factors Eomes and Tbx21 in a mouse model. As a result, it has been implicated in both the terminal maturation and functions of NK cells as well as the conversion of NK progenitor cells to immature NK cells.160,161 The administration of miR-124 by EV had synergistic anti-tumor effects by decreasing M2 microglial polarization and limiting the development of human glioblastoma cells by recruiting NK cells to the tumor.162 According to Shi et al, in BC cell lines, overexpression of miR-338-3p reduced the release of ADAM17 (a disintegrin and metalloprotease-17). Moreover, boosting granzyme B, CD16, and NKG2D production in NK cells can be achieved through the use of anti-ADAM17 antibody therapy or the overexpression of miRNA-338-3p. These educated NK cells restricted BC cell line viability. The results collectively suggest that estrogen exerts a negative influence on miR-338-3p expression in BC cells, thereby favoring the survival of these cells and compromising the function of NK cells by upregulating ADAM17, a process that ultimately hampers NK cell activity.163 Pathania et al discovered that the miR-29 family promotes the activation of NK cell immune responses in neuroblastoma (NB) by targeting the B7-H3 checkpoint. Their findings revealed that deceased patients displayed a depletion of miR-29 family members (miR-29a, miR-29b, and miR-29c), which had an inverse relationship with B7-H3 expression in NB patients. Both overexpression and knockdown studies showed that these miRNAs break down B7-H3 mRNA, which increases the cytotoxicity and stimulation of NK cells. Moreover, experiments conducted in vivo showed that members of the miR-29 family cause tumor cell apoptosis, enhance NK cell infiltration and activation, and decrease tumorigenicity, macrophage infiltration, and microvessel density.164 A recent study revealed that activated MYC in cancer cells induces a signaling pathway involving miR-29c-3p and CD276, which enables tumor cells to evade immune surveillance by suppressing the cytotoxic activity of NK cells in various types of cancer.165
Chang et al reported that treatment of NK92 cells with lung cancer cell line CL1-5 derived EVs or their transfection with miR-150-5p mimics resulted in elevated IL-10 expression and reduced CD226 expression, lytic capacity, and IFN-γ production.166 In contrast, the ability of hypoxic CL1-5 derived EVs to downregulate CD226 expression was significantly attenuated by the presence of miR-150-5p inhibitors, which in turn augmented the cytotoxic activity and IFN-γ production of NK92 cells, while suppressing IL-10 secretion.166 Exosomes derived from the bladder cancer cell line T24, have been demonstrated to contribute to the malfunction of NK cells by reducing their survival and their capacity to cytotoxically attack target cells. During this period, the expression of key functional receptors (NKG2D, NKp30, and CD226) on NK cells was significantly diminished, and their release of cytolytic proteins (granzyme-B and perforin) was also compromised by the presence of exosomes derived from T24 cells. The interference of miR-221-5p and miR-186-5p with the stability of DAP10, CD96, and the perforin gene mRNAs in NK cells has been demonstrated, and these genes may be targets for bladder cancer treatment.167 Hypoxia was found to decrease the expression of miR-1275 in NK cells as well as their cytotoxicity. The expression of the tumor suppressor axis inhibition protein (AXIN2) in NK cells is downregulated as a result of increased miR-1275 levels, which allows pancreatic cancer cells to evade the immune system.168 B7-H3 is an immunological checkpoint molecule that belongs to the B7 family. It has been demonstrated that B7-H3 controls the peripheral immunological response mediated by T cells and is linked with NK cell exhaustion in ovarian cancer. miR-29c directly targets B7-H3 in vitro, increasing the anti-tumor activity of NK cells and preventing NK cell exhaustion.169 miR-519a-3p promotes resistance to apoptosis in breast cancer cells by targeting and downregulating TRAIL-R2, caspase-8, and caspase-7, reducing their response to apoptotic signals. It also impairs NK cell-mediated tumor killing by suppressing NKG2D ligands (ULBP2 and MICA) on tumor cells.170
In addition, exosomes secreted from NK cells can also contain different miRNAs and have important effects on cancers. In this regard, it has been reported that exosomes generated from NK cells, which include miR-30c, were found to have dramatically reduced levels of miR-30c in the primary NK cells of patients with lung cancer. Overexpression of miR-30c increases TNF-α and IFN-γ production and boosts IL-2-treated NK cells' ability to kill lung cancer cells, suggesting its potential to improve NK cell cytotoxicity. Therefore, modulation of miR-30c expression could potentially enhance the cytolytic capacity of NK cells against lung cancer cells. These findings indicate that controlling the expression of miR-30c might be a potential strategy for improving the effectiveness of NK cell-based treatments against tumors.171 Another research used microarray analysis to investigate the miRNA composition of NK exosomes. Certain miRNAs, such as let-7b-5p, miR-16-5p, miR-342-3p, miR-24-3p, and miR-92a-3p, were shown to be substantially expressed in NK exosomes. Furthermore, it was shown that by specifically targeting the cell cycle regulator CDK6, NK exosomes effectively enhance let-7b-5p expression in pancreatic cancer cells and cause cell proliferation suppression.172 Based on another study, IDO1/miR-18a/NKG2D/NKG2DL axis has a key function in the regulation of NK cell function. IDO1 impairs the cytotoxicity of NK cells by suppressing NKG2D/NKG2DLs via enhancing miR-18a.173 Pathania et al demonstrated that miR-15a and miR-15b may stimulate the activation and cytotoxicity of CD8+T and NK cells against Neuroblastoma in vitro. Furthermore, the injection of murine cells that express miR-15a resulted in a decrease in tumor growth, a reduction in tumor blood vessels, and an increase in the activation and infiltration of CD8+T and NK cells into the tumors in vivo. They further demonstrated that inhibiting the expression of PD-L1 on the cell surface by employing an anti-PD-L1 antibody restored the CD8+T and NK cell-mediated anti-tumor responses elicited by miR-15a/miR-15b.174 miR-92 has higher levels of expression in ovarian cancer tissue as compared to normal cancer tissue. Transfecting miR-92 into ovarian cancer cells resulted in an increase in cell proliferation. Additionally, there was an apparent increase in migratory capability and colony formation after miR-92 transfection. Furthermore, the decrease in LATS2 (Large tumor suppressor kinase 2) expression led to the enhanced movement of YAP1 (yes-associated protein 1) and the elevation of PD-L1, which subsequently inhibited the activity of NK cells and facilitated the apoptosis of T cells.175 When activated by IL-2, NK cells displayed reduced expression of miR-301a-3p, accompanied by increased production of IFN-γ and TNF-α. Overexpression of hsa-miR-301a-3p, however, decreased the levels of these cytokines and impaired NK cell cytotoxicity. Notably, RUNX3 was identified as a target of hsa-miR-301a-3p. By inhibiting NK cells' ability to fight NSCLC cells, hsa-miR-301a-3p increased tumor development, according to in vivo studies.176 The overexpression of miR-130a enhanced the ability of NK cells to kill NSCLC cells. Further study revealed that miR-130a targets STAT3 and that the overexpression of STAT3 diminished the improvement in killing activity of NK cells against NSCLC cells induced by miR-130a. Therefore, miR-130a enhances the killing capacity of NK cells against NSCLC cells by targeting STAT3177 (Table S2, Fig. 5). One of the main obstacles to all cancer treatment strategies, including miRNA-based immunotherapy, is the heterogeneity of cancer cells. The acquisition of novel mutations may lead to heterogeneity and the formation of tumor subclones that are resistant to treatment.178 The capacity of cancer cells to change their biological state in response to therapeutic pressure, for example, by shifting into a quiescent state when exposed to drugs that kill rapidly proliferating cells, is a second aspect of heterogeneity.179 Tumor heterogeneity must thus be taken into account for improved efficacy when using miRNAs as targets for altering immune cells in the TME.
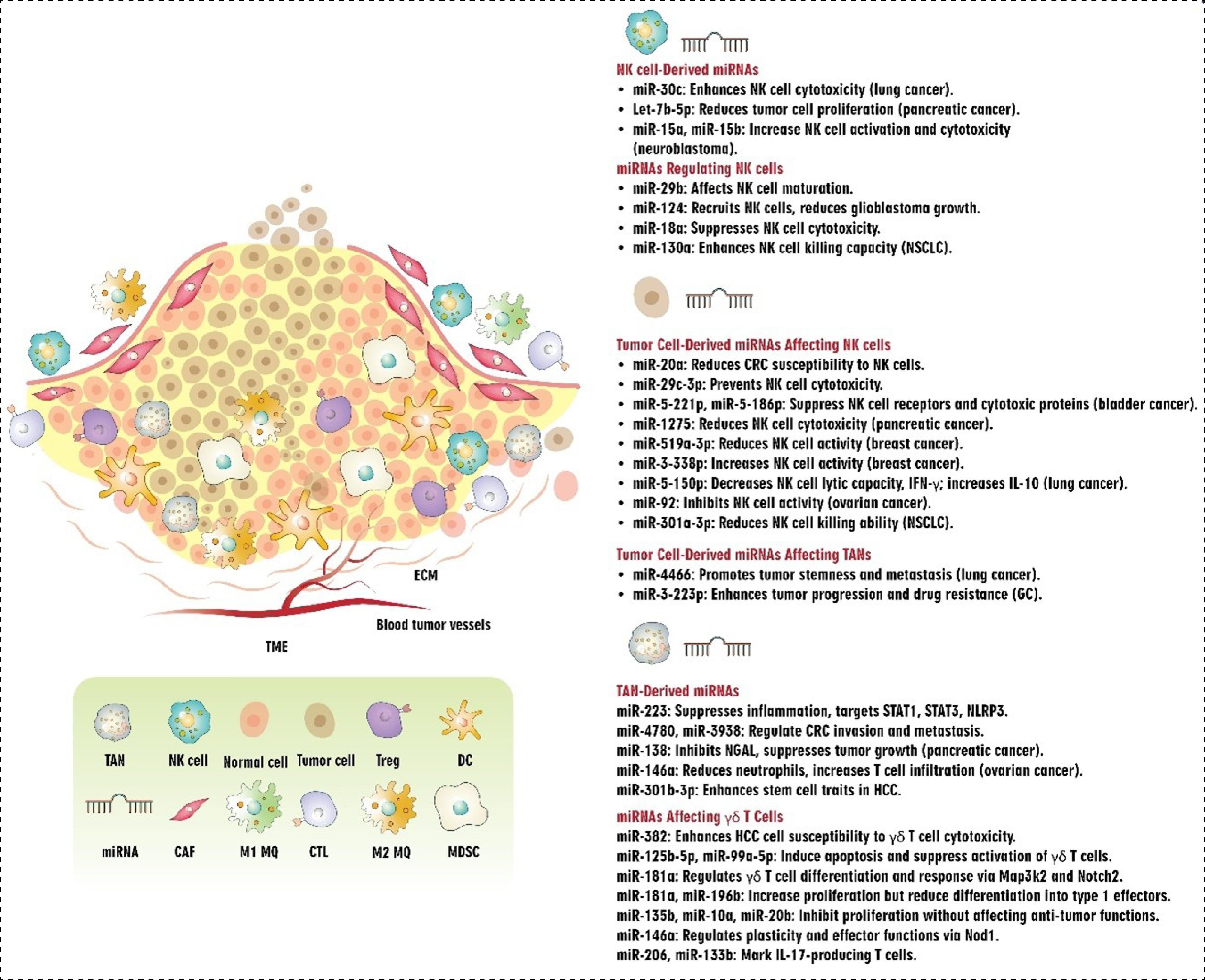
Fig. 5.
Interplay between miRNAs and NK cells/TANs/γδ T cells in TME.Abbreviations: TME: tumor microenvironment, ECM: extracellular matrix, miRNA: microRNA, TAN: tumor-associated neutrophils, NK cell: natural killer cell, Treg: T regulatory cell, DC: dendritic cell, CAF: cancer-associated fibroblast, MDSC: myeloid-derived suppressor cells, CTL: cytotoxic T cells, MQ: macrophage, NSCLC: non-small cell lung cancer, CRC: colorectal cancer, IFN: interferon, STAT: signal transducer and activator of transcription, GC: gastric cancer, NLRP3: NLR family pyrin domain containing 3, NGAL: Neutrophil Gelatinase-Associated Lipocalin, HCC: hepatocellular carcinoma.
.
Interplay between miRNAs and NK cells/TANs/γδ T cells in TME.Abbreviations: TME: tumor microenvironment, ECM: extracellular matrix, miRNA: microRNA, TAN: tumor-associated neutrophils, NK cell: natural killer cell, Treg: T regulatory cell, DC: dendritic cell, CAF: cancer-associated fibroblast, MDSC: myeloid-derived suppressor cells, CTL: cytotoxic T cells, MQ: macrophage, NSCLC: non-small cell lung cancer, CRC: colorectal cancer, IFN: interferon, STAT: signal transducer and activator of transcription, GC: gastric cancer, NLRP3: NLR family pyrin domain containing 3, NGAL: Neutrophil Gelatinase-Associated Lipocalin, HCC: hepatocellular carcinoma.
Tumor-associated neutrophils
Since neutrophils can be quickly attracted to the location of the immunological response, they are referred to as "first-line defense" cells.180 Neutrophils utilize various mechanisms, including phagocytosis, degranulation, and the formation of neutrophil extracellular traps (NETs), to mount a non-specific defense against pathogens and combat infection.180 Neutrophils have been found to possess immunoregulatory abilities, which involve the release of various substances such as cytokines and EVs. Significant biological consequences can result from even small changes in the concentration of a single miRNA molecule. miRNAs frequently aggregate into clusters, which are functional assemblages that regulate numerous constituents of a singular activity or individual constituents of interconnected pathways.181 Over 100 distinct miRNA molecules that control gene expression in immune cells have been documented.182 Human neutrophils currently exhibit 148 miRNA molecules, although this count is subject to modification when further ones are identified.182 Research has confirmed that miRNA molecules regulate every phase of immune system cell growth and function, spanning from stem cells to activated effector cells involved in both acquired and innate immune responses.183 For example, miR-130a plays a key role in the early stages of neutrophil development, where it is highly expressed and regulates the activity of crucial transcription factors such as MYB (myeloblastosis proto-oncogene) and CBF-β (core binding factor beta subunit). Moreover, it has been proposed that miR-130a may also influence the expression of proteins like myeloperoxidase (MPO) and proteinase 3, which are essential for neutrophil function.184 In addition, miR-130a also modulates the expression of the transcription factor Smad4, a key regulator of TGF-β1. Interestingly, elevated levels of miR-130a impede the normal progression of neutrophil progenitors out of the cell cycle and the development of secondary granule proteins, a process mediated by the C/EBP-ε transcription factor.185 C/EBP-ε is recognized as the primary factor that controls the final phase of neutrophil development. The synthesis of the secondary granule proteins lactoferrin, cathelicidin, and lipocalin-2 is inhibited when miR-130a is overexpressed, which results in the developing neutrophils with an immature phenotype.186 Hawez et al discovered that miR-155 plays a critical role in the formation of NETs by targeting the mRNA of peptidyl arginine deiminase 4 (PAD4), an enzyme involved in histone citrullination during NETosis.187 Moreover, Chen et al, revealed that vascular endothelial cells (VECs) treated with oxidized low-density lipoprotein (ox-LDL) that contain exosome-encapsulated miR-505 aggravate atherosclerosis via causing NET formation. They discovered that ox-LDL treatment may trigger the NF-κB pathway, which in turn triggers the transcription of miR-505. The high expression of miR-505 encapsulated in exosomes then targeted and inhibited SIRT3 in neutrophils, causing an increase in ROS levels and neutrophil NET release.188 Tumor-associated neutrophils (TANs) are cells that are present within the TME, and their roles in regulating tumor progression have been the focus of increasing research. Studies suggest that TANs have two distinct roles in this regard. The first type of TANs, known as N1 TANs, are thought to play an antitumor role, whereas N2 TANs promote tumor proliferation, metastasis, drug resistance, and recurrence by releasing various proteins, NETs, and lipids.189,190
In addition, miRNAs play a key role in regulating the differentiation of neutrophils in various cancers. In this regard, a correlation has been identified between neutrophil-derived exosomes (Neu-Exo) and the advancement of lung cancer. Exosomal miR-4466, generated by nicotine-activated N2 neutrophils, fosters tumor cell stemness and metastasis. Cancer-free individuals with a smoking history have exhibited higher levels of exosomal miR-4466 in their serum/urine, implying that miR-4466 could serve as a valuable prognostic biomarker for predicting the heightened risk of brain metastasis.191 miR-223-3p was found enriched in serum Neu-Exo and elevated in GC patients compared to benign gastric disease patients and healthy controls. Exosomal miR-223 derived from CD66b+ neutrophils is associated with tumor progression, metastasis, and drug resistance and may be a potential biomarker for cancer diagnosis and prognosis.192 Furthermore, the miR-223 molecule can have an effect on the process of neutrophil differentiation and can influence the accumulation of neutrophils in the lungs by regulating the production of CXCL2 and CCL3.193 Furthermore, exosomal miR-223 derived from neutrophils downregulates the expression of components in the canonical NF-κB pathway, thus suppressing inflammasome activation and the production of inflammatory cytokines.194 Neutrophil-derived exosomal miR-223 also directly targets STAT1, STAT3, and NLRP3 and it regulates genes involved in cancer cell proliferation, survival, differentiation, immune evasion, adhesion, and migration.194 Researchers have identified a potential link between TANs and the microenvironment of CRC. A recent study analyzed the distribution of miRNAs in N2-type neutrophils and discovered that two specific miRNAs, hsa-miR-4780 and hsa-miR-3938, were expressed at different levels compared to other types of neutrophils. Further investigation revealed that these miRNAs regulate the expression of TUSC1 and ZNF197 genes. It is thought that these miRNAs may play a role in regulating CRC invasion and metastasis by influencing the behavior of N2-type neutrophils.195 Additionally, neutrophil-derived products have been found to influence tumor growth. One such product is neutrophil gelatinase-associated lipocalin (NGAL), a small protein molecule involved in the interaction between epithelial and mesenchymal cells in tumors. The expression of NGAL in various types of cancer cells, including breast, endometrial, and pancreatic carcinomas, can be controlled by miR-138. In a study on pancreatic cancer cells, introducing miR-138 led to the suppression of NGAL, which subsequently reduced tumor cell proliferation and metastasis.196 The administration of miR-146a elicits a significant reduction in immunosuppressive neutrophil levels and a corresponding increase in CD8+T cell tumor infiltration, which is associated with a reduction in ovarian tumor burden. miR-146a targets TRAF6 and IRAK1 to inhibit NF-κB activation and decrease neutrophil infiltration into tumors through CXCL1 downregulation.197 Also, miR301b-3p plays a crucial role in enabling TANs to enhance stem cell-like properties in HCC cells. Its action specifically boosts stem cell-like traits in HCC cells and is essential to the effects induced by TANs. The interaction between TANs and miR-301b-3p leads to the activation of NF-κB signaling and an increase in CXCL5 expression.198 A recent study has made a significant discovery, revealing that neutrophil miRNA profiling may serve as a novel class of circulating biomarkers for the early detection of NSCLC. The research analyzed a dataset of 82 lung cancer patients and 73 healthy controls and identified two specific miRNAs (miRs-26a-2-3p and 574-3p) that demonstrated significant potential as diagnostic markers. These biomarkers showed an impressive sensitivity of 77.8% and specificity of 78.1% in detecting NSCLC. This finding suggests that these circulating neutrophil miRNAs may hold promise as a reliable tool for identifying lung cancer199 (Table S3, Fig. 5).
γδ T cells
γδ T cells are a unique subset of T cells with innate-like characteristics, characterized by the existence of heterodimers of T cell receptor (TCR) γ and δ.200,201 γδ T cells are abundant in peripheral tissues like the skin, lungs, and intestines. Their development in the thymus is unaffected by positive or negative selection. The unconventional innate-like T cells can quickly respond to infection and tumors via direct cytotoxicity and cytokine release after recognizing antigens in an MHC-unrestricted manner.200 γδ T cells and αβ T cells exhibit distinct characteristics in their development, distribution, proliferation patterns, and functional activities. These differences are closely linked to the miRNA-based regulatory mechanisms. While significant progress has been achieved in understanding miRNA-mediated regulation in αβ T cells, there is still a considerable knowledge gap regarding the functions of miRNAs in γδ T cells.200
Elaborating on the function of miR-382 in regulating the C-FLIP/caspase-8 pathway can provide valuable insights into its role in γδ T cell-driven cytotoxicity against HCC. Research by Chen et al. revealed that miR-382 expression is decreased in HCC tissues and cell lines.202 Elevated expression of miR-382 enhanced the susceptibility of HCC cells to γδ T cell-mediated cytotoxicity by targeting the mRNA of cellular FADD-like interleukin-1β-converting enzyme-inhibitory protein (c-FLIP). The suppression of c-FLIP by miR-382 significantly facilitated the destruction of HCC cells by boosting caspase-8 activation during γδ T cell treatment.202 c-FLIP regulates apoptosis by serving as an endogenous inhibitor of death receptor-induced apoptosis through the caspase-8 pathway. It interferes with the activation of caspase-8, thereby preventing the initiation of the extrinsic apoptotic signaling cascade.203 miR-382 reduces the inhibitory action of c-FLIP, allowing caspase-8 to be activated. Once activated, caspase-8 cleaves and triggers downstream effector caspases, such as caspase-3, ultimately resulting in apoptosis of cancer cells. This mechanism highlights the therapeutic potential of miR-382 in enhancing the sensitivity of HCC cells to immune cell-mediated cytotoxicity.204 Given the need to investigate how miRNAs regulate circulating human γδ T cells, a study was conducted on healthy donors. This study analyzed miRNA expression patterns in peripheral γδ T cells. There were 14 miRNAs expressed differently by peripheral αβ T cells and γδ T cells. Subsequent investigations into these miRNAs validated that miR-125b-5p and miR-99a-5p induce apoptosis in γδ T cells and suppress their activation, diminishing their cytotoxicity to tumor cells.205 miR-181a has been identified as a novel molecular regulator of functional differentiation in human γδ T cells. It has been demonstrated that both its -5p and -2-3p strands regulate T cells' differentiation and responsiveness towards type 1 effectors by targeting Map3k2 and Notch2 mRNAs. Map3k2, a kinase involved in the MAPK signaling pathway, plays a crucial role in regulating inflammatory responses. By targeting Map3k2, miR-181a modulates the production of pro-inflammatory cytokines and influences γδ T cell responses within the tumor microenvironment.206 Notch2, a key regulator in the Notch signaling pathway, is essential for T cell differentiation and lineage commitment. Through the suppression of Notch2 expression, miR-181a alters the functional plasticity of γδ T cells, impacting their anti-tumor activities.207 These regulatory mechanisms are finely tuned by the TME's inflammatory and immunosuppressive cues, highlighting the potential of miR-181a as a therapeutic target for modulating γδ T cell functions in cancer.208
T cells are a critical part of the immune system. They target a wide variety of malignant cells with their cytolytic and inflammatory properties. In clinical trials, only 10-33% of patients have shown objective responses. Thus, it is evident that to manipulate γδ T cells in clinical settings, it is vital to understand the mechanisms involved in regulating their activation and functional differentiation.209 In this regard, Gordino et al observation revealed that an increase in γδ T cell proliferation occurred simultaneously with a decrease in their differentiation into type 1 effectors. This was due to miR-181a and miR-196b overexpression. Additionally, miR-135b, miR-10a, and miR-20b were found to impede γδ T cell proliferation in their preliminary findings, without affecting their anti-tumor functions. If confirmed, these findings could have significant implications for γδ T cell manipulation in cancer immunotherapy.209 Recent studies have elucidated how miR-146a modulates γδ T cell functions by targeting Nod1, an intracellular pattern recognition receptor involved in immune signaling. miR-146a is highly expressed in γδ27⁻ T cells, a subset preprogrammed to produce IL-17.210 This microRNA acts as a post-transcriptional regulator, limiting the production of IFN-γ in these cells.211 By targeting Nod1 mRNA, miR-146a reduces Nod1 protein levels, thereby influencing γδ T cell plasticity, the ability of these cells to adapt their functional profile in response to environmental cues.210 Nod1 plays a crucial role in promoting IFN-γ production in γδ27⁻ T cells. In the absence of miR-146a, increased Nod1 expression leads to the emergence of γδ T cells capable of producing both IL-17 and IFN-γ, indicating enhanced functional plasticity.211 Conversely, Nod1-deficient mice exhibit a lack of multifunctional IL-17⁺ IFN-γ⁺ γδ27⁻ T cells and show increased susceptibility to infections, such as Listeria monocytogenes, highlighting the importance of Nod1 in effective immune responses.210,212 The miR-146a/Nod1 axis also affects crosstalk with other immune cells. γδ T cells influence the activation and differentiation of various immune cell types, including macrophages and DCs through modulating cytokine production.211 For instance, the balance between IL-17 and IFN-γ production can shape the inflammatory milieu, altering the recruitment and activation of innate immune cells, which in turn modulate adaptive immune responses.210,212 The co-regulation of neighboring genes is a complex process influenced by various factors. Researchers have discovered that miR-206/133b and IL-17A are both inducible by IL-23 signaling, implying that shared regulatory elements may contribute to their coordinated regulation. The activation of STAT3 by IL-23/p19 leads to direct binding of phosphorylated STAT3 to the promoters of IL-17A and IL-17F, suggesting that both chromatin remodeling and shared regulatory elements may play a role in the coordinated regulation of miR-206/133b and IL-17A.213 The recent finding highlights the intriguing phenomenon of co-regulation between miR-133b and miR-206 with the Il17a/F locus, a feature that is unique to T cell differentiation. Interestingly, this characteristic is conserved across mouse αβ and γδ T cells, as well as human Th17 cells. As a result, miR-133b and miR-206 have been identified as potential biomarkers for T cells that produce IL-17.213 (Fig. 5)
Tumor heterogeneity significantly influences the miRNAs' involvement in immune regulation. For instance, miR-382 has been shown to sensitize HCC cells to γδ T cell-mediated cytotoxicity by targeting c-FLIP.202 In breast cancer, miR-382 has been linked to tumor invasion by targeting distinct pathways, demonstrating the context-dependent nature of miRNA functions.214 Similarly, miR-181a, which regulates γδ T cell plasticity through Map3k2 and Notch2 and promotes prostate cancer progression, demonstrates varying expression levels and effects in different tumor microenvironments, such as HCC, gastric and glioblastoma cancers.206,215-217 miR-181a-5p is downregulated in HCC and inversely correlated with Egr1 expression. Functional studies reveal that miR-181a-5p overexpression suppresses HCC cell proliferation, while its suppression enhances proliferation. Mechanistically, miR-181a-5p directly targets Egr1, leading to the downregulation of the TGF-β1/Smad pathway.215 In gastric cancer, miR-181a acts as a negative regulator of autophagy in cisplatin-resistant SGC7901/CDDP cells by targeting ATG5. Overexpression of miR-181a enhances cisplatin sensitivity in vitro and reduces tumor growth in gastric cancer xenografts.216 miR-181a replacement in glioblastoma cells increases sensitivity to low-dose carmustine by promoting apoptosis, reducing viability, and causing cell cycle arrest. It regulates pro-apoptotic and anti-apoptotic genes, suppresses metastasis via MMP-2 and Bach1 downregulation, and inhibits proliferation through AKT1 suppression, implicating the PI3K/AKT pathway.217 These differences highlight the importance of considering tumor-specific characteristics when designing miRNA-based therapies. Although the regulation of tumor-associated innate immune cells by miRNAs is the main emphasis of this study, miRNAs also affect adaptive immunity. For example, miR-155 enhances T helper cell differentiation by targeting SOCS1, thus promoting anti-tumor immunity.218 On the other hand, miR-150 regulates B cell maturation and antibody production, indirectly influencing tumor progression.219 Although it is still little understood, the interaction of miRNAs, γδ T cells, and adaptive immune cells shows promise for further research, particularly in the context of combination treatments that target both innate and adaptive immune responses.
Future perspectives on miRNA-based therapeutics
miRNA-based therapeutics hold a bright future in cancer treatment; however, translation from bench to bedside is still the most significant challenge, including suitable administration methods, stability within the body, tissue- and cell- specific targeting, and achieving the desired intracellular effects.220,221 Another complication during miRNA therapy is hypoxia in cancerous tissues, causing downregulation of Drosha, Dicer, and AGO2, the main enzymes in biogenesis and proper actions of miRNAs.222 Thus, only a limited array of miRNA therapeutics has progressed into clinical trials. Currently, two strategies mainly have been employed in miRNA therapy.223 One approach is miRNA restoration therapy, which uses synthetic oligonucleotides (miRNA mimics) to replace the downregulated or non-functional miRNA. The other approach is miRNA inhibition therapy, which aims to silence the overexpressed miRNA by utilizing antagonists (anti-miRs).223,224 The most common route for the administration of therapeutic miRNA is via either intravenous or intracutaneous injection.225 Meanwhile, researchers are developing new routes for administering miRNA-based therapies. One of these routes involves using a biodegradable 3D matrix containing miRNA therapeutics after removing the affected tissue during surgery.226,227 Another new approach that has attracted researchers' attention is the oral administration of plant-derived miRNAs. However, this route requires further exploration regarding its bioavailability and functionality.228
Some modifications could potentially improve miRNA stability in the in vivo environments. One strategy is to replace the 2’-OH moieties with 2’-O-methyl (2’-O-Me) or 2’-fluoro (2’-F) substituents, reducing toxicity and increasing target binding.229 Conjugating mimics with cholesterol is another way to increase cellular uptakes.230 Another approach is substituting the phosphodiester with phosphorothioate (PS) linkages, which cause evading intracellular nucleases.229 Conjugation of mimics and antitumor drugs is also reported as an effective approach; combining miR-15a and miR-194 mimics with Gemcitabine showed higher efficacy compared to the sole use of each one, both in vitro and in vivo models of PDAC.231
Several clinical trials have been started focusing on miRNAs in various conditions. However, they could encounter some complications. The best instance is MRX34, miR-34a mimic, in a cancer treatment clinical trial (NCT01829971), which induced immune-related side effects and caused the death of four patients, which put an end to the trial.232,233 This miRNA was delivered systemically with a liposomal amphoteric strategy, which should be taken up by low PH cancerous tissue.234 However, pre-clinical experiments showed the deployment of this miRNA in the spleen and bone marrow, suggesting immune-related complication.235,236 RG-101, an anti-miR-122 drug, was another therapeutic under investigation at the phase II clinical trial for treating chronic hepatitis C virus (HCV). Unfortunately, it has been terminated due to the induction of hyperbilirubinemia in several patients during treatment.237
It is clear that miRNA-based therapies are still in the early clinical stages. Meanwhile, TRYNGOLZA, Lumasiran, Givosiran, Patisiran, and Inclisiran are the number of FDA-approved siRNA products, indicating the advancement in RNA-based therapies in the clinic. However, altering the miRNA levels in humans can result in physiological disruption and unknown reactions. Therefore, more investigation into the side effects of miRNAs is crucial, but it is not unlikely that miRNAs will receive FDA approval for clinical use in the near future.
Conclusion
This study provides new insights into the intricate interactions among tumor-released miRNAs and the innate immune system within the TME. Unlike the prevalent focus on adaptive immunity in current literature, the manuscript highlights the dual functions of miRNAs in modulating immunosuppression and immunostimulation through innate immune cells, such as macrophages, dendritic cells, MDSCs, and NK cells. One of the main arguments made is that miRNAs play a context-dependent and dynamic role—some miRNAs promote tumor progression through M2 polarization or by expanding MDSCs, while others enhance anti-tumor responses through M1 polarization or by reactivating dendritic and NK cells.
Additionally, the research highlights the basic importance of the transport of miRNAs through extracellular vesicles, which allows for bidirectional communication between immune and tumor cells. This process is central to the establishment of feedback loops that maintain tumor-supportive microenvironments, thus further elucidating complexities in the framework of immune evasion. Notably, the comprehensive examination of regulatory processes mediated through miRNAs in diverse types of cancers presents great hopes for both therapy and diagnosis, particularly in regards to modulation of innate immune cell functions and bypassing drug resistance. Overall, the findings present opportunities for miRNA-directed strategies to transform the immune microenvironment within tumors and maximize the effectiveness of cancer immunotherapy.
Review Highlights
What is the current knowledge?
-
TME shapes cancer progression through complex interactions with immune and stromal cells.
-
Innate immune cells can both promote and inhibit tumor growth, influencing immunotherapy outcomes.
-
Tumor-derived miRNAs regulate gene expression in immune cells and contribute to immune evasion and drug resistance.
What is new here?
-
Our study highlights the dual roles of miRNAs in creating either immunosuppressive or immunogenic environments.
-
The study suggests miRNA-innate immune cell interactions as novel targets to enhance cancer immunotherapy.
Competing Interests
The authors have no competing interests to declare.
Ethical Approval
Not applicable.
Supplementary files
Supplementary file 1 contains Tables S1-S3.
(pdf)
References
- Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell 2011; 144:646-74. doi: 10.1016/j.cell.2011.02.013 [Crossref] [ Google Scholar]
- Elhanani O, Ben-Uri R, Keren L. Spatial profiling technologies illuminate the tumor microenvironment. Cancer Cell 2023; 41:404-20. doi: 10.1016/j.ccell.2023.01.010 [Crossref] [ Google Scholar]
- de Visser KE, Joyce JA. The evolving tumor microenvironment: from cancer initiation to metastatic outgrowth. Cancer Cell 2023; 41:374-403. doi: 10.1016/j.ccell.2023.02.016 [Crossref] [ Google Scholar]
- Lu C, Liu Y, Ali NM, Zhang B, Cui X. The role of innate immune cells in the tumor microenvironment and research progress in anti-tumor therapy. Front Immunol 2022; 13:1039260. doi: 10.3389/fimmu.2022.1039260 [Crossref] [ Google Scholar]
- Leone P, Malerba E, Susca N, Favoino E, Perosa F, Brunori G. Endothelial cells in tumor microenvironment: insights and perspectives. Front Immunol 2024; 15:1367875. doi: 10.3389/fimmu.2024.1367875 [Crossref] [ Google Scholar]
- Hinshaw DC, Shevde LA. The tumor microenvironment innately modulates cancer progression. Cancer Res 2019; 79:4557-66. doi: 10.1158/0008-5472.Can-18-3962 [Crossref] [ Google Scholar]
- Poltavets V, Kochetkova M, Pitson SM, Samuel MS. The role of the extracellular matrix and its molecular and cellular regulators in cancer cell plasticity. Front Oncol 2018; 8:431. doi: 10.3389/fonc.2018.00431 [Crossref] [ Google Scholar]
- Brabletz T, Pfeuffer I, Schorr E, Siebelt F, Wirth T, Serfling E. Transforming growth factor beta and cyclosporin A inhibit the inducible activity of the interleukin-2 gene in T cells through a noncanonical octamer-binding site. Mol Cell Biol 1993; 13:1155-62. doi: 10.1128/mcb.13.2.1155-1162.1993 [Crossref] [ Google Scholar]
- Patel H, Nilendu P, Jahagirdar D, Pal JK, Sharma NK. Modulating secreted components of tumor microenvironment: a masterstroke in tumor therapeutics. Cancer Biol Ther 2018; 19:3-12. doi: 10.1080/15384047.2017.1394538 [Crossref] [ Google Scholar]
- Postow MA, Callahan MK, Wolchok JD. Immune checkpoint blockade in cancer therapy. J Clin Oncol 2015; 33:1974-82. doi: 10.1200/jco.2014.59.4358 [Crossref] [ Google Scholar]
- Alizadeh AA, Aranda V, Bardelli A, Blanpain C, Bock C, Borowski C. Toward understanding and exploiting tumor heterogeneity. Nat Med 2015; 21:846-53. doi: 10.1038/nm.3915 [Crossref] [ Google Scholar]
- Proietto M, Crippa M, Damiani C, Pasquale V, Sacco E, Vanoni M. Tumor heterogeneity: preclinical models, emerging technologies, and future applications. Front Oncol 2023; 13:1164535. doi: 10.3389/fonc.2023.1164535 [Crossref] [ Google Scholar]
- Li F, Simon MC. Cancer cells don't live alone: metabolic communication within tumor microenvironments. Dev Cell 2020; 54:183-95. doi: 10.1016/j.devcel.2020.06.018 [Crossref] [ Google Scholar]
- Anderson NM, Simon MC. The tumor microenvironment. Curr Biol 2020; 30:R921-5. doi: 10.1016/j.cub.2020.06.081 [Crossref] [ Google Scholar]
- Gajewski TF, Schreiber H, Fu YX. Innate and adaptive immune cells in the tumor microenvironment. Nat Immunol 2013; 14:1014-22. doi: 10.1038/ni.2703 [Crossref] [ Google Scholar]
- Binnewies M, Roberts EW, Kersten K, Chan V, Fearon DF, Merad M. Understanding the tumor immune microenvironment (TIME) for effective therapy. Nat Med 2018; 24:541-50. doi: 10.1038/s41591-018-0014-x [Crossref] [ Google Scholar]
- Rajbhandary S, Dhakal H, Shrestha S. Tumor immune microenvironment (TIME) to enhance antitumor immunity. Eur J Med Res 2023; 28:169. doi: 10.1186/s40001-023-01125-3 [Crossref] [ Google Scholar]
- Sadeghi M, Dehnavi S, Sharifat M, Amiri AM, Khodadadi A. Innate immune cells: Key players of orchestra in modulating tumor microenvironment (TME). Heliyon 2024; 10:e27480. doi: 10.1016/j.heliyon.2024.e27480 [Crossref] [ Google Scholar]
- Philip M, Schietinger A. CD8 + T cell differentiation and dysfunction in cancer. Nat Rev Immunol 2022; 22:209-23. doi: 10.1038/s41577-021-00574-3 [Crossref] [ Google Scholar]
- Borst J, Ahrends T, Bąbała N, Melief CJ, Kastenmüller W. CD4 + T cell help in cancer immunology and immunotherapy. Nat Rev Immunol 2018; 18:635-47. doi: 10.1038/s41577-018-0044-0 [Crossref] [ Google Scholar]
- Togashi Y, Shitara K, Nishikawa H. Regulatory T cells in cancer immunosuppression - implications for anticancer therapy. Nat Rev Clin Oncol 2019; 16:356-71. doi: 10.1038/s41571-019-0175-7 [Crossref] [ Google Scholar]
- Laumont CM, Banville AC, Gilardi M, Hollern DP, Nelson BH. Tumour-infiltrating B cells: immunological mechanisms, clinical impact and therapeutic opportunities. Nat Rev Cancer 2022; 22:414-30. doi: 10.1038/s41568-022-00466-1 [Crossref] [ Google Scholar]
- Moghaddasnejad MR, Keshavarz A, Mardi A, Sherafat NS, Aghebati-Maleki L, Mohammadi MH. LncRNAs as behind-the-scenes molecules in cancer progression through regulating tumor-associated innate immune system cells. Mol Biol Rep 2025; 52:449. doi: 10.1007/s11033-025-10513-3 [Crossref] [ Google Scholar]
- Pagès F, Galon J, Dieu-Nosjean MC, Tartour E, Sautès-Fridman C, Fridman WH. Immune infiltration in human tumors: a prognostic factor that should not be ignored. Oncogene 2010; 29:1093-102. doi: 10.1038/onc.2009.416 [Crossref] [ Google Scholar]
- Doedens AL, Stockmann C, Rubinstein MP, Liao D, Zhang N, DeNardo DG. Macrophage expression of hypoxia-inducible factor-1 alpha suppresses T-cell function and promotes tumor progression. Cancer Res 2010; 70:7465-75. doi: 10.1158/0008-5472.Can-10-1439 [Crossref] [ Google Scholar]
- Fridlender ZG, Sun J, Kim S, Kapoor V, Cheng G, Ling L. Polarization of tumor-associated neutrophil phenotype by TGF-beta: "N1" versus "N2" TAN. Cancer Cell 2009; 16:183-94. doi: 10.1016/j.ccr.2009.06.017 [Crossref] [ Google Scholar]
- Wculek SK, Malanchi I. Neutrophils support lung colonization of metastasis-initiating breast cancer cells. Nature 2015; 528:413-7. doi: 10.1038/nature16140 [Crossref] [ Google Scholar]
- Linde IL, Prestwood TR, Qiu J, Pilarowski G, Linde MH, Zhang X, et al. Neutrophil-activating therapy for the treatment of cancer. Cancer Cell 2023; 41: 356-72.e10. doi: 10.1016/j.ccell.2023.01.002.
- Ni J, Wang X, Stojanovic A, Zhang Q, Wincher M, Bühler L, et al. Single-cell RNA sequencing of tumor-infiltrating NK cells reveals that inhibition of transcription factor HIF-1α unleashes NK cell activity. Immunity 2020; 52: 1075-87.e8. doi: 10.1016/j.immuni.2020.05.001.
- Cao H, Gao S, Jogani R, Sugimura R. The tumor microenvironment reprograms immune cells. Cell Reprogram 2022; 24:343-52. doi: 10.1089/cell.2022.0047 [Crossref] [ Google Scholar]
- Schoenfeld DA, Moutafi M, Martinez S, Djureinovic D, Merkin RD, Adeniran A. Immune dysfunction revealed by digital spatial profiling of immuno-oncology markers in progressive stages of renal cell carcinoma and in brain metastases. J Immunother Cancer 2023; 11:e007240. doi: 10.1136/jitc-2023-007240 [Crossref] [ Google Scholar]
- Cong J, Wang X, Zheng X, Wang D, Fu B, Sun R, et al. Dysfunction of natural killer cells by FBP1-induced inhibition of glycolysis during lung cancer progression. Cell Metab 2018; 28: 243-55.e5. doi: 10.1016/j.cmet.2018.06.021.
- Marzagalli M, Ebelt ND, Manuel ER. Unraveling the crosstalk between melanoma and immune cells in the tumor microenvironment. Semin Cancer Biol 2019; 59:236-50. doi: 10.1016/j.semcancer.2019.08.002 [Crossref] [ Google Scholar]
- Wu CJ, Lu LF. MicroRNA in immune regulation. Curr Top Microbiol Immunol 2017; 410:249-67. doi: 10.1007/82_2017_65 [Crossref] [ Google Scholar]
- Gebert LF, MacRae IJ. Regulation of microRNA function in animals. Nat Rev Mol Cell Biol 2019; 20:21-37. doi: 10.1038/s41580-018-0045-7 [Crossref] [ Google Scholar]
- Annese T, Tamma R, De Giorgis M, Ribatti D. microRNAs biogenesis, functions and role in tumor angiogenesis. Front Oncol 2020; 10:581007. doi: 10.3389/fonc.2020.581007 [Crossref] [ Google Scholar]
- Wang WT, Han C, Sun YM, Chen TQ, Chen YQ. Noncoding RNAs in cancer therapy resistance and targeted drug development. J Hematol Oncol 2019; 12:55. doi: 10.1186/s13045-019-0748-z [Crossref] [ Google Scholar]
- Chen X, Ying X, Wang X, Wu X, Zhu Q, Wang X. Exosomes derived from hypoxic epithelial ovarian cancer deliver microRNA-940 to induce macrophage M2 polarization. Oncol Rep 2017; 38:522-8. doi: 10.3892/or.2017.5697 [Crossref] [ Google Scholar]
- Yoshimura A, Sawada K, Nakamura K, Kinose Y, Nakatsuka E, Kobayashi M. Exosomal miR-99a-5p is elevated in sera of ovarian cancer patients and promotes cancer cell invasion by increasing fibronectin and vitronectin expression in neighboring peritoneal mesothelial cells. BMC Cancer 2018; 18:1065. doi: 10.1186/s12885-018-4974-5 [Crossref] [ Google Scholar]
- Sánchez-Sendra B, García-Giménez JL, González-Muñoz JF, Navarro L, Murgui A, Terrádez L. Circulating miRNA expression analysis reveals new potential biomarkers for human cutaneous melanoma staging. J Eur Acad Dermatol Venereol 2020; 34:e126-e9. doi: 10.1111/jdv.16060 [Crossref] [ Google Scholar]
- Sabry D, El-Deek SE, Maher M, El-Baz MA, El-Bader HM, Amer E. Role of miRNA-210, miRNA-21 and miRNA-126 as diagnostic biomarkers in colorectal carcinoma: impact of HIF-1α-VEGF signaling pathway. Mol Cell Biochem 2019; 454:177-89. doi: 10.1007/s11010-018-3462-1 [Crossref] [ Google Scholar]
- Basak U, Sarkar T, Mukherjee S, Chakraborty S, Dutta A, Dutta S. Tumor-associated macrophages: an effective player of the tumor microenvironment. Front Immunol 2023; 14:1295257. doi: 10.3389/fimmu.2023.1295257 [Crossref] [ Google Scholar]
- Pan Y, Yu Y, Wang X, Zhang T. Tumor-associated macrophages in tumor immunity. Front Immunol 2020; 11:583084. doi: 10.3389/fimmu.2020.583084 [Crossref] [ Google Scholar]
- Chen Y, Jin H, Song Y, Huang T, Cao J, Tang Q. Targeting tumor-associated macrophages: a potential treatment for solid tumors. J Cell Physiol 2021; 236:3445-65. doi: 10.1002/jcp.30139 [Crossref] [ Google Scholar]
- Moeini P, Niedźwiedzka-Rystwej P. Tumor-associated macrophages: combination of therapies, the approach to improve cancer treatment. Int J Mol Sci 2021; 22:7239. doi: 10.3390/ijms22137239 [Crossref] [ Google Scholar]
- Tan Y, Wang M, Zhang Y, Ge S, Zhong F, Xia G. Tumor-associated macrophages: a potential target for cancer therapy. Front Oncol 2021; 11:693517. doi: 10.3389/fonc.2021.693517 [Crossref] [ Google Scholar]
- Su P, Li O, Ke K, Jiang Z, Wu J, Wang Y. Targeting tumor-associated macrophages: critical players in tumor progression and therapeutic strategies (review). Int J Oncol 2024; 64:60. doi: 10.3892/ijo.2024.5648 [Crossref] [ Google Scholar]
- Cheng Y, Song S, Wu P, Lyu B, Qin M, Sun Y. Tumor associated macrophages and TAMs-based anti-tumor nanomedicines. Adv Healthc Mater 2021; 10:e2100590. doi: 10.1002/adhm.202100590 [Crossref] [ Google Scholar]
- Boutilier AJ, Elsawa SF. Macrophage polarization states in the tumor microenvironment. Int J Mol Sci 2021; 22:6995. doi: 10.3390/ijms22136995 [Crossref] [ Google Scholar]
- Mills CD, Kincaid K, Alt JM, Heilman MJ, Hill AM. M-1/M-2 macrophages and the Th1/Th2 paradigm. J Immunol 2000; 164:6166-73. doi: 10.4049/jimmunol.164.12.6166 [Crossref] [ Google Scholar]
- Orecchioni M, Ghosheh Y, Pramod AB, Ley K. Macrophage polarization: different gene signatures in M1(LPS + ) vs classically and M2(LPS-) vs alternatively activated macrophages. Front Immunol 2019; 10:1084. doi: 10.3389/fimmu.2019.01084 [Crossref] [ Google Scholar]
- Qin R, Ren W, Ya G, Wang B, He J, Ren S. Role of chemokines in the crosstalk between tumor and tumor-associated macrophages. Clin Exp Med 2023; 23:1359-73. doi: 10.1007/s10238-022-00888-z [Crossref] [ Google Scholar]
- Sezginer O, Unver N. Dissection of pro-tumoral macrophage subtypes and immunosuppressive cells participating in M2 polarization. Inflamm Res 2024; 73:1411-23. doi: 10.1007/s00011-024-01907-3 [Crossref] [ Google Scholar]
- Zhang Q, Sioud M. Tumor-associated macrophage subsets: shaping polarization and targeting. Int J Mol Sci 2023; 24:7493. doi: 10.3390/ijms24087493 [Crossref] [ Google Scholar]
- Mohapatra S, Pioppini C, Ozpolat B, Calin GA. Non-coding RNAs regulation of macrophage polarization in cancer. Mol Cancer 2021; 20:24. doi: 10.1186/s12943-021-01313-x [Crossref] [ Google Scholar]
- Wang C, Wang X, Zhang D, Sun X, Wu Y, Wang J. The macrophage polarization by miRNAs and its potential role in the treatment of tumor and inflammation (review). Oncol Rep 2023; 50:190. doi: 10.3892/or.2023.8627 [Crossref] [ Google Scholar]
- Curtale G, Rubino M, Locati M. MicroRNAs as molecular switches in macrophage activation. Front Immunol 2019; 10:799. doi: 10.3389/fimmu.2019.00799 [Crossref] [ Google Scholar]
- Khayati S, Dehnavi S, Sadeghi M, Tavakol Afshari J, Esmaeili SA, Mohammadi M. The potential role of miRNA in regulating macrophage polarization. Heliyon 2023; 9:e21615. doi: 10.1016/j.heliyon.2023.e21615 [Crossref] [ Google Scholar]
- Zhou X, Chen B, Zhang Z, Huang Y, Li J, Wei Q. Crosstalk between tumor-associated macrophages and microRNAs: a key role in tumor microenvironment. Int J Mol Sci 2022; 23:13258. doi: 10.3390/ijms232113258 [Crossref] [ Google Scholar]
- Yu MY, Jia HJ, Zhang J, Ran GH, Liu Y, Yang XH. Exosomal miRNAs-mediated macrophage polarization and its potential clinical application. Int Immunopharmacol 2023; 117:109905. doi: 10.1016/j.intimp.2023.109905 [Crossref] [ Google Scholar]
- Kwon Y, Kim M, Kim Y, Jung HS, Jeoung D. Exosomal microRNAs as mediators of cellular interactions between cancer cells and macrophages. Front Immunol 2020; 11:1167. doi: 10.3389/fimmu.2020.01167 [Crossref] [ Google Scholar]
- Li L, Sun P, Zhang C, Li Z, Cui K, Zhou W. MiR-98 modulates macrophage polarization and suppresses the effects of tumor-associated macrophages on promoting invasion and epithelial-mesenchymal transition of hepatocellular carcinoma. Cancer Cell Int 2018; 18:95. doi: 10.1186/s12935-018-0590-3 [Crossref] [ Google Scholar]
- Wang L, Hu YY, Zhao JL, Huang F, Liang SQ, Dong L, et al. Targeted delivery of miR-99b reprograms tumor-associated macrophage phenotype leading to tumor regression. J Immunother Cancer 2020; 8. doi: 10.1136/jitc-2019-000517.
- Zhao J, Li H, Zhao S, Wang E, Zhu J, Feng D. Epigenetic silencing of miR-144/451a cluster contributes to HCC progression via paracrine HGF/MIF-mediated TAM remodeling. Mol Cancer 2021; 20:46. doi: 10.1186/s12943-021-01343-5 [Crossref] [ Google Scholar]
- Ma D, Zhang Y, Chen G, Yan J. miR-148a affects polarization of THP-1-derived macrophages and reduces recruitment of tumor-associated macrophages via targeting SIRPα. Cancer Manag Res 2020; 12:8067-77. doi: 10.2147/cmar.S238317 [Crossref] [ Google Scholar]
- Zhong C, Tao B, Yang F, Xia K, Yang X, Chen L. Histone demethylase JMJD1C promotes the polarization of M1 macrophages to prevent glioma by upregulating miR-302a. Clin Transl Med 2021; 11:e424. doi: 10.1002/ctm2.424 [Crossref] [ Google Scholar]
- Moradi-Chaleshtori M, Bandehpour M, Heidari N, Mohammadi-Yeganeh S, Mahmoud Hashemi S. Exosome-mediated miR-33 transfer induces M1 polarization in mouse macrophages and exerts antitumor effect in 4T1 breast cancer cell line. Int Immunopharmacol 2021; 90:107198. doi: 10.1016/j.intimp.2020.107198 [Crossref] [ Google Scholar]
- Yang T, Han Y, Chen J, Liang X, Sun L. MiR-506 promotes antitumor immune response in pancreatic cancer by reprogramming tumor-associated macrophages toward an M1 phenotype. Biomedicines 2023; 11:2874. doi: 10.3390/biomedicines11112874 [Crossref] [ Google Scholar]
- Zhang LS, Chen QC, Zong HT, Xia Q. Exosome miRNA-203 promotes M1 macrophage polarization and inhibits prostate cancer tumor progression. Mol Cell Biochem 2024; 479:2459-70. doi: 10.1007/s11010-023-04854-5 [Crossref] [ Google Scholar]
- Zhao S, Mi Y, Guan B, Zheng B, Wei P, Gu Y. Tumor-derived exosomal miR-934 induces macrophage M2 polarization to promote liver metastasis of colorectal cancer. J Hematol Oncol 2020; 13:156. doi: 10.1186/s13045-020-00991-2 [Crossref] [ Google Scholar]
- Yin Y, Liu B, Cao Y, Yao S, Liu Y, Jin G. Colorectal cancer-derived small extracellular vesicles promote tumor immune evasion by upregulating PD-L1 expression in tumor-associated macrophages. Adv Sci (Weinh) 2022; 9:2102620. doi: 10.1002/advs.202102620 [Crossref] [ Google Scholar]
- Shinohara H, Kuranaga Y, Kumazaki M, Sugito N, Yoshikawa Y, Takai T. Regulated polarization of tumor-associated macrophages by miR-145 via colorectal cancer-derived extracellular vesicles. J Immunol 2017; 199:1505-15. doi: 10.4049/jimmunol.1700167 [Crossref] [ Google Scholar]
- Li M, Xu H, Qi Y, Pan Z, Li B, Gao Z. Tumor-derived exosomes deliver the tumor suppressor miR-3591-3p to induce M2 macrophage polarization and promote glioma progression. Oncogene 2022; 41:4618-32. doi: 10.1038/s41388-022-02457-w [Crossref] [ Google Scholar]
- Chen J, Zhang K, Zhi Y, Wu Y, Chen B, Bai J. Tumor-derived exosomal miR-19b-3p facilitates M2 macrophage polarization and exosomal LINC00273 secretion to promote lung adenocarcinoma metastasis via Hippo pathway. Clin Transl Med 2021; 11:e478. doi: 10.1002/ctm2.478 [Crossref] [ Google Scholar]
- Ma J, Chen S, Liu Y, Han H, Gong M, Song Y. The role of exosomal miR-181b in the crosstalk between NSCLC cells and tumor-associated macrophages. Genes Genomics 2022; 44:1243-58. doi: 10.1007/s13258-022-01227-y [Crossref] [ Google Scholar]
- Ma C, He D, Tian P, Wang Y, He Y, Wu Q. miR-182 targeting reprograms tumor-associated macrophages and limits breast cancer progression. Proc Natl Acad Sci U S A 2022; 119:e2114006119. doi: 10.1073/pnas.2114006119 [Crossref] [ Google Scholar]
- Xun J, Du L, Gao R, Shen L, Wang D, Kang L. Cancer-derived exosomal miR-138-5p modulates polarization of tumor-associated macrophages through inhibition of KDM6B. Theranostics 2021; 11:6847-59. doi: 10.7150/thno.51864 [Crossref] [ Google Scholar]
- Cobos Jiménez V, Bradley EJ, Willemsen AM, van Kampen AH, Baas F, Kootstra NA. Next-generation sequencing of microRNAs uncovers expression signatures in polarized macrophages. Physiol Genomics 2014; 46:91-103. doi: 10.1152/physiolgenomics.00140.2013 [Crossref] [ Google Scholar]
- Ito I, Asai A, Suzuki S, Kobayashi M, Suzuki F. M2b macrophage polarization accompanied with reduction of long noncoding RNA GAS5. Biochem Biophys Res Commun 2017; 493:170-5. doi: 10.1016/j.bbrc.2017.09.053 [Crossref] [ Google Scholar]
- Sánchez-González I, Bobien A, Molnar C, Schmid S, Strotbek M, Boerries M. miR-149 suppresses breast cancer metastasis by blocking paracrine interactions with macrophages. Cancer Res 2020; 80:1330-41. doi: 10.1158/0008-5472.Can-19-1934 [Crossref] [ Google Scholar]
- Hao C, Sheng Z, Wang W, Feng R, Zheng Y, Xiao Q. Tumor-derived exosomal miR-148b-3p mediates M2 macrophage polarization via TSC2/mTORC1 to promote breast cancer migration and invasion. Thorac Cancer 2023; 14:1477-91. doi: 10.1111/1759-7714.14891 [Crossref] [ Google Scholar]
- Ni J, Xi X, Xiao S, Xiao X. Tumor cell-derived exosomal miR-191-5p activates M2-subtype macrophages through SOCS3 to facilitate breast cancer. Mol Biotechnol 2024. 66: 1314-25. doi: 10.1007/s12033-023-01034-0.
- Zhou H, Gan M, Jin X, Dai M, Wang Y, Lei Y. miR-382 inhibits breast cancer progression and metastasis by affecting the M2 polarization of tumor-associated macrophages by targeting PGC-1α. Int J Oncol 2022; 61:126. doi: 10.3892/ijo.2022.5416 [Crossref] [ Google Scholar]
- Qiu S, Xie L, Lu C, Gu C, Xia Y, Lv J. Gastric cancer-derived exosomal miR-519a-3p promotes liver metastasis by inducing intrahepatic M2-like macrophage-mediated angiogenesis. J Exp Clin Cancer Res 2022; 41:296. doi: 10.1186/s13046-022-02499-8 [Crossref] [ Google Scholar]
- Ye Z, Yi J, Jiang X, Shi W, Xu H, Cao H. Gastric cancer-derived exosomal let-7 g-5p mediated by SERPINE1 promotes macrophage M2 polarization and gastric cancer progression. J Exp Clin Cancer Res 2025; 44:2. doi: 10.1186/s13046-024-03269-4 [Crossref] [ Google Scholar]
- Yang C, Dou R, Wei C, Liu K, Shi D, Zhang C. Tumor-derived exosomal microRNA-106b-5p activates EMT-cancer cell and M2-subtype TAM interaction to facilitate CRC metastasis. Mol Ther 2021; 29:2088-107. doi: 10.1016/j.ymthe.2021.02.006 [Crossref] [ Google Scholar]
- Pei W, Wei K, Wu Y, Qiu Q, Zhu H, Mao L. Colorectal cancer tumor cell-derived exosomal miR-203a-3p promotes CRC metastasis by targeting PTEN-induced macrophage polarization. Gene 2023; 885:147692. doi: 10.1016/j.gene.2023.147692 [Crossref] [ Google Scholar]
- Shi X, Wei K, Wu Y, Mao L, Pei W, Zhu H. Exosome-derived miR-372-5p promotes stemness and metastatic ability of CRC cells by inducing macrophage polarization. Cell Signal 2023; 111:110884. doi: 10.1016/j.cellsig.2023.110884 [Crossref] [ Google Scholar]
- Bagheri V, Khorramdelazad H, Hassanshahi G, Moghadam-Ahmadi A, Vakilian A. CXCL12 and CXCR4 in the peripheral blood of patients with Parkinson's disease. Neuroimmunomodulation 2018; 25:201-5. doi: 10.1159/000494435 [Crossref] [ Google Scholar]
- Azin H, Vazirinejad R, Nasiri Ahmadabadi B, Khorramdelazad H, Rezazadeh Zarandi E, Kazemi Arababadi M. The SDF-1 3'a genetic variation of the chemokine SDF-1α (CXCL12) in parallel with its increased circulating levels is associated with susceptibility to MS: a study on Iranian multiple sclerosis patients. J Mol Neurosci 2012; 47:431-6. doi: 10.1007/s12031-011-9672-6 [Crossref] [ Google Scholar]
- Hassanshahi G, Shamsizadeh A, Kazemi Arababadi M, Khoramdelazad H, Yaghini N, Rezazadeh Zarandi E. Assessment of CXCL12 (SDF-1α) polymorphisms and its serum level in posttransfusion occult HBV-infected patients in Southeastern Iran. Int J Infect Dis 2012; 16:e86. doi: 10.1016/j.ijid.2012.05.200 [Crossref] [ Google Scholar]
- Aminzadeh F, Ghorashi Z, Nabati S, Ghasemshirazi M, Kazemi Arababadi M, Shamsizadeh A. Differential expression of CXC chemokines CXCL 10 and CXCL 12 in term and pre‐term neonates and their mothers. Am J Reprod Immunol 2012; 68:338-44. doi: 10.1111/j.1600-0897.2012.01167.x [Crossref] [ Google Scholar]
- Khorramdelazad H, Bagheri V, Hassanshahi G, Zeinali M, Vakilian A. New insights into the role of stromal cell-derived factor 1 (SDF-1/CXCL12) in the pathophysiology of multiple sclerosis. J Neuroimmunol 2016; 290:70-5. doi: 10.1016/j.jneuroim.2015.11.021 [Crossref] [ Google Scholar]
- Nazari A, Khorramdelazad H, Hassanshahi G. Biological/pathological functions of the CXCL12/CXCR4/CXCR7 axes in the pathogenesis of bladder cancer. Int J Clin Oncol 2017; 22:991-1000. doi: 10.1007/s10147-017-1187-x [Crossref] [ Google Scholar]
- Chen J, Li Z, Yue C, Ma J, Cao L, Lin J. Human umbilical cord mesenchymal stem cell-derived exosomes carrying miR-1827 downregulate SUCNR1 to inhibit macrophage M2 polarization and prevent colorectal liver metastasis. Apoptosis 2023; 28:549-65. doi: 10.1007/s10495-022-01798-x [Crossref] [ Google Scholar]
- Xu Y, Luan G, Liu F, Zhang Y, Li Z, Liu Z. Exosomal miR-200b-3p induce macrophage polarization by regulating transcriptional repressor ZEB1 in hepatocellular carcinoma. Hepatol Int 2023; 17:889-903. doi: 10.1007/s12072-023-10507-y [Crossref] [ Google Scholar]
- Wang Y, Ge WL, Wang SJ, Liu YY, Zhang ZH, Hua Y. MiR-548t-5p regulates pancreatic ductal adenocarcinoma metastasis through an IL-33-dependent crosstalk between cancer cells and M2 macrophages. Cell Cycle 2024; 23:169-87. doi: 10.1080/15384101.2024.2309026 [Crossref] [ Google Scholar]
- Liu J, Luo R, Wang J, Luan X, Wu D, Chen H. Tumor cell-derived exosomal miR-770 inhibits M2 macrophage polarization via targeting MAP3K1 to inhibit the invasion of non-small cell lung cancer cells. Front Cell Dev Biol 2021; 9:679658. doi: 10.3389/fcell.2021.679658 [Crossref] [ Google Scholar]
- Zhang C, Chen W, Pan S, Zhang S, Xie H, Zhang Z. SEVs-mediated miR-6750 transfer inhibits pre-metastatic niche formation in nasopharyngeal carcinoma by targeting M6PR. Cell Death Discov 2023; 9:2. doi: 10.1038/s41420-022-01262-4 [Crossref] [ Google Scholar]
- Chen Y, Lei Y, Li J, Wang X, Li G. Macrophage-derived exosomal microRNAs promote metastasis in pancreatic ductal adenocarcinoma. Int Immunopharmacol 2024; 129:111590. doi: 10.1016/j.intimp.2024.111590 [Crossref] [ Google Scholar]
- Yin Z, Ma T, Huang B, Lin L, Zhou Y, Yan J. Macrophage-derived exosomal microRNA-501-3p promotes progression of pancreatic ductal adenocarcinoma through the TGFBR3-mediated TGF-β signaling pathway. J Exp Clin Cancer Res 2019; 38:310. doi: 10.1186/s13046-019-1313-x [Crossref] [ Google Scholar]
- Liu Q, Zhao E, Geng B, Gao S, Yu H, He X. Tumor-associated macrophage-derived exosomes transmitting miR-193a-5p promote the progression of renal cell carcinoma via TIMP2-dependent vasculogenic mimicry. Cell Death Dis 2022; 13:382. doi: 10.1038/s41419-022-04814-9 [Crossref] [ Google Scholar]
- Feng J, Xu B, Dai C, Wang Y, Xie G, Yang W. Macrophage-derived exosomal miR-342-3p promotes the progression of renal cell carcinoma through the NEDD4L/CEP55 axis. Oncol Res 2021; 29:331-49. doi: 10.32604/or.2022.03554 [Crossref] [ Google Scholar]
- Yang X, Cai S, Shu Y, Deng X, Zhang Y, He N. Exosomal miR-487a derived from m2 macrophage promotes the progression of gastric cancer. Cell Cycle 2021; 20:434-44. doi: 10.1080/15384101.2021.1878326 [Crossref] [ Google Scholar]
- Zhang R, Chen X, Miao C, Chen Y, Li Y, Shen J. Tumor-associated macrophage-derived exosomal miR-513b-5p is a target of Jianpi Yangzheng decoction for inhibiting gastric cancer. J Ethnopharmacol 2024; 318:117013. doi: 10.1016/j.jep.2023.117013 [Crossref] [ Google Scholar]
- Mi X, Xu R, Hong S, Xu T, Zhang W, Liu M. M2 macrophage-derived exosomal lncRNA AFAP1-AS1 and microRNA-26a affect cell migration and metastasis in esophageal cancer. Mol Ther Nucleic Acids 2020; 22:779-90. doi: 10.1016/j.omtn.2020.09.035 [Crossref] [ Google Scholar]
- Peng J, Li S, Li B, Hu W, Ding C. Exosomes derived from M1 macrophages inhibit the proliferation of the A549 and H1299 lung cancer cell lines via the miRNA-let-7b-5p-GNG5 axis. PeerJ 2023; 11:e14608. doi: 10.7717/peerj.14608 [Crossref] [ Google Scholar]
- Zheng P, Chen L, Yuan X, Luo Q, Liu Y, Xie G. Exosomal transfer of tumor-associated macrophage-derived miR-21 confers cisplatin resistance in gastric cancer cells. J Exp Clin Cancer Res 2017; 36:53. doi: 10.1186/s13046-017-0528-y [Crossref] [ Google Scholar]
- An Y, Yang Q. MiR-21 modulates the polarization of macrophages and increases the effects of M2 macrophages on promoting the chemoresistance of ovarian cancer. Life Sci 2020; 242:117162. doi: 10.1016/j.lfs.2019.117162 [Crossref] [ Google Scholar]
- Zhu X, Shen H, Yin X, Yang M, Wei H, Chen Q. Macrophages derived exosomes deliver miR-223 to epithelial ovarian cancer cells to elicit a chemoresistant phenotype. J Exp Clin Cancer Res 2019; 38:81. doi: 10.1186/s13046-019-1095-1 [Crossref] [ Google Scholar]
- Gao H, Ma J, Cheng Y, Zheng P. Exosomal transfer of macrophage-derived miR-223 confers doxorubicin resistance in gastric cancer. Onco Targets Ther 2020; 13:12169-79. doi: 10.2147/ott.S283542 [Crossref] [ Google Scholar]
- Wang H, Wang L, Pan H, Wang Y, Shi M, Yu H. Exosomes derived from macrophages enhance aerobic glycolysis and chemoresistance in lung cancer by stabilizing c-Myc via the inhibition of NEDD4L. Front Cell Dev Biol 2020; 8:620603. doi: 10.3389/fcell.2020.620603 [Crossref] [ Google Scholar]
- Guo Y, Wu H, Xiong J, Gou S, Cui J, Peng T. miR-222-3p-containing macrophage-derived extracellular vesicles confer gemcitabine resistance via TSC1-mediated mTOR/AKT/PI3K pathway in pancreatic cancer. Cell Biol Toxicol 2023; 39:1203-14. doi: 10.1007/s10565-022-09736-y [Crossref] [ Google Scholar]
- Guo X, Qiu W, Liu Q, Qian M, Wang S, Zhang Z. Immunosuppressive effects of hypoxia-induced glioma exosomes through myeloid-derived suppressor cells via the miR-10a/Rora and miR-21/PTEN pathways. Oncogene 2018; 37:4239-59. doi: 10.1038/s41388-018-0261-9 [Crossref] [ Google Scholar]
- Guo X, Qiu W, Wang J, Liu Q, Qian M, Wang S. Glioma exosomes mediate the expansion and function of myeloid-derived suppressor cells through microRNA-29a/Hbp1 and microRNA-92a/Prkar1a pathways. Int J Cancer 2019; 144:3111-26. doi: 10.1002/ijc.32052 [Crossref] [ Google Scholar]
- Qiu W, Guo X, Li B, Wang J, Qi Y, Chen Z. Exosomal miR-1246 from glioma patient body fluids drives the differentiation and activation of myeloid-derived suppressor cells. Mol Ther 2021; 29:3449-64. doi: 10.1016/j.ymthe.2021.06.023 [Crossref] [ Google Scholar]
- Li L, Cao B, Liang X, Lu S, Luo H, Wang Z. Microenvironmental oxygen pressure orchestrates an anti- and pro-tumoral γδ T cell equilibrium via tumor-derived exosomes. Oncogene 2019; 38:2830-43. doi: 10.1038/s41388-018-0627-z [Crossref] [ Google Scholar]
- Hu X, Li J, Fu M, Zhao X, Wang W. The JAK/STAT signaling pathway: from bench to clinic. Signal Transduct Target Ther 2021; 6:402. doi: 10.1038/s41392-021-00791-1 [Crossref] [ Google Scholar]
- Jiang M, Zhang W, Zhang R, Liu P, Ye Y, Yu W. Cancer exosome-derived miR-9 and miR-181a promote the development of early-stage MDSCs via interfering with SOCS3 and PIAS3 respectively in breast cancer. Oncogene 2020; 39:4681-94. doi: 10.1038/s41388-020-1322-4 [Crossref] [ Google Scholar]
- Bruns H, Böttcher M, Qorraj M, Fabri M, Jitschin S, Dindorf J. CLL-cell-mediated MDSC induction by exosomal miR-155 transfer is disrupted by vitamin D. Leukemia 2017; 31:985-8. doi: 10.1038/leu.2016.378 [Crossref] [ Google Scholar]
- Ren W, Zhang X, Li W, Feng Q, Feng H, Tong Y. Exosomal miRNA-107 induces myeloid-derived suppressor cell expansion in gastric cancer. Cancer Manag Res 2019; 11:4023-40. doi: 10.2147/cmar.S198886 [Crossref] [ Google Scholar]
- Basso D, Gnatta E, Padoan A, Fogar P, Furlanello S, Aita A. PDAC-derived exosomes enrich the microenvironment in MDSCs in a SMAD4-dependent manner through a new calcium related axis. Oncotarget 2017; 8:84928-44. doi: 10.18632/oncotarget.20863 [Crossref] [ Google Scholar]
- Zhou JH, Yao ZX, Zheng Z, Yang J, Wang R, Fu SJ. G-MDSCs-derived exosomal miRNA-143-3p promotes proliferation via targeting of ITM2B in lung cancer. Onco Targets Ther 2020; 13:9701-19. doi: 10.2147/ott.S256378 [Crossref] [ Google Scholar]
- Mackert JR, Qu P, Min Y, Johnson PF, Yang L, Lin PC. Dual negative roles of C/EBPα in the expansion and pro-tumor functions of MDSCs. Sci Rep 2017; 7:14048. doi: 10.1038/s41598-017-12968-2 [Crossref] [ Google Scholar]
- Jiang J, Gao Q, Gong Y, Huang L, Lin H, Zhou X. MiR-486 promotes proliferation and suppresses apoptosis in myeloid cells by targeting Cebpa in vitro. Cancer Med 2018; 7:4627-38. doi: 10.1002/cam4.1694 [Crossref] [ Google Scholar]
- Sun JP, Ge QX, Ren Z, Sun XF, Xie SP. MiR-6991-3p is identified as a novel suppressor in the expansion and activation of myeloid-derived suppressor cells in hepatoma-bearing mice. Onco Targets Ther 2019; 12:309-17. doi: 10.2147/ott.S185422 [Crossref] [ Google Scholar]
- Lv Y, Ma X, Ma Y, Du Y, Feng J. A new emerging target in cancer immunotherapy: galectin-9 (LGALS9). Genes Dis 2023; 10:2366-82. doi: 10.1016/j.gendis.2022.05.020 [Crossref] [ Google Scholar]
- Mevel R, Draper JE, Lie-A-Ling M, Kouskoff V, Lacaud G. RUNX transcription factors: orchestrators of development. Development 2019; 146:dev148296. doi: 10.1242/dev.148296 [Crossref] [ Google Scholar]
- Tian J, Rui K, Tang X, Ma J, Wang Y, Tian X. MicroRNA-9 regulates the differentiation and function of myeloid-derived suppressor cells via targeting Runx1. J Immunol 2015; 195:1301-11. doi: 10.4049/jimmunol.1500209 [Crossref] [ Google Scholar]
- Xu Z, Ji J, Xu J, Li D, Shi G, Liu F. MiR-30a increases MDSC differentiation and immunosuppressive function by targeting SOCS3 in mice with B-cell lymphoma. FEBS J 2017; 284:2410-24. doi: 10.1111/febs.14133 [Crossref] [ Google Scholar]
- Xin P, Xu X, Deng C, Liu S, Wang Y, Zhou X. The role of JAK/STAT signaling pathway and its inhibitors in diseases. Int Immunopharmacol 2020; 80:106210. doi: 10.1016/j.intimp.2020.106210 [Crossref] [ Google Scholar]
- Chen S, Wang L, Fan J, Ye C, Dominguez D, Zhang Y. Host miR155 promotes tumor growth through a myeloid-derived suppressor cell-dependent mechanism. Cancer Res 2015; 75:519-31. doi: 10.1158/0008-5472.Can-14-2331 [Crossref] [ Google Scholar]
- Li L, Zhang J, Diao W, Wang D, Wei Y, Zhang CY. MicroRNA-155 and microRNA-21 promote the expansion of functional myeloid-derived suppressor cells. J Immunol 2014; 192:1034-43. doi: 10.4049/jimmunol.1301309 [Crossref] [ Google Scholar]
- Wang J, Yu F, Jia X, Iwanowycz S, Wang Y, Huang S. MicroRNA-155 deficiency enhances the recruitment and functions of myeloid-derived suppressor cells in tumor microenvironment and promotes solid tumor growth. Int J Cancer 2015; 136:E602-13. doi: 10.1002/ijc.29151 [Crossref] [ Google Scholar]
- Mei S, Xin J, Liu Y, Zhang Y, Liang X, Su X. MicroRNA-200c promotes suppressive potential of myeloid-derived suppressor cells by modulating PTEN and FOG2 expression. PLoS One 2015; 10:e0135867. doi: 10.1371/journal.pone.0135867 [Crossref] [ Google Scholar]
- Liu Y, Lai L, Chen Q, Song Y, Xu S, Ma F. MicroRNA-494 is required for the accumulation and functions of tumor-expanded myeloid-derived suppressor cells via targeting of PTEN. J Immunol 2012; 188:5500-10. doi: 10.4049/jimmunol.1103505 [Crossref] [ Google Scholar]
- Diao J, Gu H, Tang M, Zhao J, Cattral MS. Tumor dendritic cells (DCs) derived from precursors of conventional DCs are dispensable for intratumor CTL responses. J Immunol 2018; 201:1306-14. doi: 10.4049/jimmunol.1701514 [Crossref] [ Google Scholar]
- Stumpfova Z, Hezova R, Meli AC, Slaby O, Michalek J. MicroRNA profiling of activated and tolerogenic human dendritic cells. Mediators Inflamm 2014; 2014:259689. doi: 10.1155/2014/259689 [Crossref] [ Google Scholar]
- Ding G, Zhou L, Qian Y, Fu M, Chen J, Chen J. Pancreatic cancer-derived exosomes transfer miRNAs to dendritic cells and inhibit RFXAP expression via miR-212-3p. Oncotarget 2015; 6:29877-88. doi: 10.18632/oncotarget.4924 [Crossref] [ Google Scholar]
- Fu Y, Li F, Zhang P, Liu M, Qian L, Lv F. Myrothecine A modulates the proliferation of HCC cells and the maturation of dendritic cells through downregulating miR-221. Int Immunopharmacol 2019; 75:105783. doi: 10.1016/j.intimp.2019.105783 [Crossref] [ Google Scholar]
- Gao X, Han D, Fan W. Down-regulation of RBP-J mediated by microRNA-133a suppresses dendritic cells and functions as a potential tumor suppressor in osteosarcoma. Exp Cell Res 2016; 349:264-72. doi: 10.1016/j.yexcr.2016.10.019 [Crossref] [ Google Scholar]
- Dunand-Sauthier I, Santiago-Raber ML, Capponi L, Vejnar CE, Schaad O, Irla M. Silencing of c-Fos expression by microRNA-155 is critical for dendritic cell maturation and function. Blood 2011; 117:4490-500. doi: 10.1182/blood-2010-09-308064 [Crossref] [ Google Scholar]
- Dunand-Sauthier I, Irla M, Carnesecchi S, Seguín-Estévez Q, Vejnar CE, Zdobnov EM. Repression of arginase-2 expression in dendritic cells by microRNA-155 is critical for promoting T cell proliferation. J Immunol 2014; 193:1690-700. doi: 10.4049/jimmunol.1301913 [Crossref] [ Google Scholar]
- Wang J, Iwanowycz S, Yu F, Jia X, Leng S, Wang Y. microRNA-155 deficiency impairs dendritic cell function in breast cancer. Oncoimmunology 2016; 5:e1232223. doi: 10.1080/2162402x.2016.1232223 [Crossref] [ Google Scholar]
- Cui ZJ, Xie XL, Qi W, Yang YC, Bai Y, Han J. Cell-free miR-17-5p as a diagnostic biomarker for gastric cancer inhibits dendritic cell maturation. Onco Targets Ther 2019; 12:2661-75. doi: 10.2147/ott.S197682 [Crossref] [ Google Scholar]
- Zhou M, Chen J, Zhou L, Chen W, Ding G, Cao L. Pancreatic cancer derived exosomes regulate the expression of TLR4 in dendritic cells via miR-203. Cell Immunol 2014; 292:65-9. doi: 10.1016/j.cellimm.2014.09.004 [Crossref] [ Google Scholar]
- Tittarelli A, Navarrete M, Lizana M, Hofmann-Vega F, Salazar-Onfray F. Hypoxic melanoma cells deliver microRNAs to dendritic cells and cytotoxic T lymphocytes through connexin-43 channels. Int J Mol Sci 2020; 21:7567. doi: 10.3390/ijms21207567 [Crossref] [ Google Scholar]
- Jia J, Li X, Guo S, Xie X. MicroRNA-155 suppresses the translation of p38 and impairs the functioning of dendritic cells in endometrial cancer mice. Cancer Manag Res 2020; 12:2993-3002. doi: 10.2147/cmar.S240926 [Crossref] [ Google Scholar]
- Liang X, Liu Y, Mei S, Zhang M, Xin J, Zhang Y. MicroRNA-22 impairs anti-tumor ability of dendritic cells by targeting p38. PLoS One 2015; 10:e0121510. doi: 10.1371/journal.pone.0121510 [Crossref] [ Google Scholar]
- Guo J, Jin H, Xi Y, Guo J, Jin Y, Jiang D. The miR-582/CD1B axis is involved in regulation of dendritic cells and is associated with clinical outcomes in advanced lung adenocarcinoma. Biomed Res Int 2020; 2020:4360930. doi: 10.1155/2020/4360930 [Crossref] [ Google Scholar]
- Hsu YL, Chen YJ, Chang WA, Jian SF, Fan HL, Wang JY. Interaction between tumor-associated dendritic cells and colon cancer cells contributes to tumor progression via CXCL1. Int J Mol Sci 2018; 19:2427. doi: 10.3390/ijms19082427 [Crossref] [ Google Scholar]
- Wang J, Ye H, Zhang D, Cheng K, Hu Y, Yu X. Cancer-derived circulating microRNAs promote tumor angiogenesis by entering dendritic cells to degrade highly complementary microRNAs. Theranostics 2017; 7:1407-21. doi: 10.7150/thno.18262 [Crossref] [ Google Scholar]
- Pyfferoen L, Brabants E, Everaert C, De Cabooter N, Heyns K, Deswarte K. The transcriptome of lung tumor-infiltrating dendritic cells reveals a tumor-supporting phenotype and a microRNA signature with negative impact on clinical outcome. Oncoimmunology 2017; 6:e1253655. doi: 10.1080/2162402x.2016.1253655 [Crossref] [ Google Scholar]
- Paul S, Lal G. The molecular mechanism of natural killer cells function and its importance in cancer immunotherapy. Front Immunol 2017; 8:1124. doi: 10.3389/fimmu.2017.01124 [Crossref] [ Google Scholar]
- Liu H, Wang S, Xin J, Wang J, Yao C, Zhang Z. Role of NKG2D and its ligands in cancer immunotherapy. Am J Cancer Res 2019; 9:2064-78. [ Google Scholar]
- Guerra N, Tan YX, Joncker NT, Choy A, Gallardo F, Xiong N. NKG2D-deficient mice are defective in tumor surveillance in models of spontaneous malignancy. Immunity 2008; 28:571-80. doi: 10.1016/j.immuni.2008.02.016 [Crossref] [ Google Scholar]
- Liu YF, Chiang Y, Hsu FM, Tsai CL, Cheng JC. Radiosensitization effect by HDAC inhibition improves NKG2D-dependent natural killer cytotoxicity in hepatocellular carcinoma. Front Oncol 2022; 12:1009089. doi: 10.3389/fonc.2022.1009089 [Crossref] [ Google Scholar]
- Tang S, Fu H, Xu Q, Zhou Y. miR-20a regulates sensitivity of colorectal cancer cells to NK cells by targeting MICA. Biosci Rep 2019; 39:BSR20180695. doi: 10.1042/bsr20180695 [Crossref] [ Google Scholar]
- Youlin K, Simin L, Jian K, Li Z. Inhibition of miR-20a by pterostilbene facilitates prostate cancer cells killed by NK cells via up-regulation of NKG2D ligands and TGF-β1down-regulation. Heliyon 2023; 9:e14957. doi: 10.1016/j.heliyon.2023.e14957 [Crossref] [ Google Scholar]
- Mundy-Bosse BL, Scoville SD, Chen L, McConnell K, Mao HC, Ahmed EH. MicroRNA-29b mediates altered innate immune development in acute leukemia. J Clin Invest 2016; 126:4404-16. doi: 10.1172/jci85413 [Crossref] [ Google Scholar]
- Qi Y, Huang Y, Pang L, Gu W, Wang N, Hu J. Prognostic value of the microRNA-29 family in multiple human cancers: a meta-analysis and systematic review. Clin Exp Pharmacol Physiol 2017; 44:441-54. doi: 10.1111/1440-1681.12726 [Crossref] [ Google Scholar]
- Hong S, You JY, Paek K, Park J, Kang SJ, Han EH. Inhibition of tumor progression and M2 microglial polarization by extracellular vesicle-mediated microRNA-124 in a 3D microfluidic glioblastoma microenvironment. Theranostics 2021; 11:9687-704. doi: 10.7150/thno.60851 [Crossref] [ Google Scholar]
- Shi Y, Pan J, Hang C, Tan L, Hu L, Yan Z. The estrogen/miR-338-3p/ADAM17 axis enhances the viability of breast cancer cells via suppressing NK cell's function. Environ Toxicol 2023; 38:1618-27. doi: 10.1002/tox.23791 [Crossref] [ Google Scholar]
- Pathania AS, Chava H, Chaturvedi NK, Chava S, Byrareddy SN, Coulter DW. The miR-29 family facilitates the activation of NK-cell immune responses by targeting the B7-H3 immune checkpoint in neuroblastoma. Cell Death Dis 2024; 15:428. doi: 10.1038/s41419-024-06791-7 [Crossref] [ Google Scholar]
- Lee CC, Ho KH, Huang TW, Shih CM, Hsu SY, Liu AJ. A regulatory loop among CD276, miR-29c-3p, and Myc exists in cancer cells against natural killer cell cytotoxicity. Life Sci 2021; 277:119438. doi: 10.1016/j.lfs.2021.119438 [Crossref] [ Google Scholar]
- Chang WA, Tsai MJ, Hung JY, Wu KL, Tsai YM, Huang YC. miR-150-5p-containing extracellular vesicles are a new immunoregulator that favor the progression of lung cancer in hypoxic microenvironments by altering the phenotype of NK cells. Cancers (Basel) 2021; 13:6252. doi: 10.3390/cancers13246252 [Crossref] [ Google Scholar]
- Huyan T, Gao L, Gao N, Wang C, Guo W, Zhou X. miR-221-5p and miR-186-5p are the critical bladder cancer derived exosomal miRNAs in natural killer cell dysfunction. Int J Mol Sci 2022; 23:15177. doi: 10.3390/ijms232315177 [Crossref] [ Google Scholar]
- Ou Z, Lu Y, Xu D, Luo Z. Hypoxia mediates immune escape of pancreatic cancer cells by affecting miR-1275/AXIN2 in natural killer cells. Front Immunol 2023; 14:1271603. doi: 10.3389/fimmu.2023.1271603 [Crossref] [ Google Scholar]
- Deng M, Wu D, Zhang Y, Jin Z, Miao J. MiR-29c downregulates tumor-expressed B7-H3 to mediate the antitumor NK-cell functions in ovarian cancer. Gynecol Oncol 2021; 162:190-9. doi: 10.1016/j.ygyno.2021.04.013 [Crossref] [ Google Scholar]
- Breunig C, Pahl J, Küblbeck M, Miller M, Antonelli D, Erdem N. MicroRNA-519a-3p mediates apoptosis resistance in breast cancer cells and their escape from recognition by natural killer cells. Cell Death Dis 2017; 8:e2973. doi: 10.1038/cddis.2017.364 [Crossref] [ Google Scholar]
- Gao F, Han J, Jia L, He J, Wang Y, Chen M. MiR-30c facilitates natural killer cell cytotoxicity to lung cancer through targeting GALNT7. Genes Genomics 2023; 45:247-60. doi: 10.1007/s13258-022-01306-0 [Crossref] [ Google Scholar]
- Di Pace AL, Pelosi A, Fiore PF, Tumino N, Besi F, Quatrini L. MicroRNA analysis of Natural Killer cell-derived exosomes: the microRNA let-7b-5p is enriched in exosomes and participates in their anti-tumor effects against pancreatic cancer cells. Oncoimmunology 2023; 12:2221081. doi: 10.1080/2162402x.2023.2221081 [Crossref] [ Google Scholar]
- Zhang J, Han X, Hu X, Jin F, Gao Z, Yin L. IDO1 impairs NK cell cytotoxicity by decreasing NKG2D/NKG2DLs via promoting miR-18a. Mol Immunol 2018; 103:144-55. doi: 10.1016/j.molimm.2018.09.011 [Crossref] [ Google Scholar]
- Pathania AS, Prathipati P, Olwenyi OA, Chava S, Smith OV, Gupta SC. miR-15a and miR-15b modulate natural killer and CD8 + T-cell activation and anti-tumor immune response by targeting PD-L1 in neuroblastoma. Mol Ther Oncolytics 2022; 25:308-29. doi: 10.1016/j.omto.2022.03.010 [Crossref] [ Google Scholar]
- Feng S, Sun H, Zhu W. MiR-92 overexpression suppresses immune cell function in ovarian cancer via LATS2/YAP1/PD-L1 pathway. Clin Transl Oncol 2021; 23:450-8. doi: 10.1007/s12094-020-02439-y [Crossref] [ Google Scholar]
- Zhang J, Yang Y, Wei Y, Li L, Wang X, Ye Z. Hsa-miR-301a-3p inhibited the killing effect of natural killer cells on non-small cell lung cancer cells by regulating RUNX3. Cancer Biomark 2023; 37:249-59. doi: 10.3233/cbm-220469 [Crossref] [ Google Scholar]
- Zhou X, Liu S, Liu J, Zhang Z, Mao X, Zhou H. MicroRNA-130a enhances the killing ability of natural killer cells against non-small cell lung cancer cells by targeting signal transducers and activators of transcription 3. Biochem Biophys Res Commun 2020; 523:481-6. doi: 10.1016/j.bbrc.2019.11.099 [Crossref] [ Google Scholar]
- Vitale I, Shema E, Loi S, Galluzzi L. Intratumoral heterogeneity in cancer progression and response to immunotherapy. Nat Med 2021; 27:212-24. doi: 10.1038/s41591-021-01233-9 [Crossref] [ Google Scholar]
- Kyrysyuk O, Wucherpfennig KW. Designing cancer immunotherapies that engage T cells and NK cells. Annu Rev Immunol 2023; 41:17-38. doi: 10.1146/annurev-immunol-101921-044122 [Crossref] [ Google Scholar]
- Kraus RF, Gruber MA. Neutrophils-from bone marrow to first-line defense of the innate immune system. Front Immunol 2021; 12:767175. doi: 10.3389/fimmu.2021.767175 [Crossref] [ Google Scholar]
- Shang R, Lee S, Senavirathne G, Lai EC. microRNAs in action: biogenesis, function and regulation. Nat Rev Genet 2023; 24:816-33. doi: 10.1038/s41576-023-00611-y [Crossref] [ Google Scholar]
- Garley M, Nowak K, Jabłońska E. Neutrophil microRNAs. Biol Rev Camb Philos Soc 2024; 99:864-77. doi: 10.1111/brv.13048 [Crossref] [ Google Scholar]
- Jung N, Schenten V, Bueb JL, Tolle F, Bréchard S. miRNAs regulate cytokine secretion induced by phosphorylated S100A8/A9 in neutrophils. Int J Mol Sci 2019; 20:5699. doi: 10.3390/ijms20225699 [Crossref] [ Google Scholar]
- Pedersen CC, Refsgaard JC, Østergaard O, Jensen LJ, Heegaard NH, Borregaard N. Impact of microRNA-130a on the neutrophil proteome. BMC Immunol 2015; 16:70. doi: 10.1186/s12865-015-0134-8 [Crossref] [ Google Scholar]
- Larsen MT, Hother C, Häger M, Pedersen CC, Theilgaard-Mönch K, Borregaard N. MicroRNA profiling in human neutrophils during bone marrow granulopoiesis and in vivo exudation. PLoS One 2013; 8:e58454. doi: 10.1371/journal.pone.0058454 [Crossref] [ Google Scholar]
- Häger M, Pedersen CC, Larsen MT, Andersen MK, Hother C, Grønbæk K. MicroRNA-130a-mediated down-regulation of Smad4 contributes to reduced sensitivity to TGF-β1 stimulation in granulocytic precursors. Blood 2011; 118:6649-59. doi: 10.1182/blood-2011-03-339978 [Crossref] [ Google Scholar]
- Hawez A, Al-Haidari A, Madhi R, Rahman M, Thorlacius H. MiR-155 regulates PAD4-dependent formation of neutrophil extracellular traps. Front Immunol 2019; 10:2462. doi: 10.3389/fimmu.2019.02462 [Crossref] [ Google Scholar]
- Chen L, Hu L, Li Q, Ma J, Li H. Exosome-encapsulated miR-505 from ox-LDL-treated vascular endothelial cells aggravates atherosclerosis by inducing NET formation. Acta Biochim Biophys Sin (Shanghai) 2019; 51:1233-41. doi: 10.1093/abbs/gmz123 [Crossref] [ Google Scholar]
- Tian S, Chu Y, Hu J, Ding X, Liu Z, Fu D. Tumour-associated neutrophils secrete AGR2 to promote colorectal cancer metastasis via its receptor CD98hc-xCT. Gut 2022; 71:2489-501. doi: 10.1136/gutjnl-2021-325137 [Crossref] [ Google Scholar]
- Perego M, Tyurin VA, Tyurina YY, Yellets J, Nacarelli T, Lin C. Reactivation of dormant tumor cells by modified lipids derived from stress-activated neutrophils. Sci Transl Med 2020; 12:eabb5817. doi: 10.1126/scitranslmed.abb5817 [Crossref] [ Google Scholar]
- Tyagi A, Wu SY, Sharma S, Wu K, Zhao D, Deshpande R. Exosomal miR-4466 from nicotine-activated neutrophils promotes tumor cell stemness and metabolism in lung cancer metastasis. Oncogene 2022; 41:3079-92. doi: 10.1038/s41388-022-02322-w [Crossref] [ Google Scholar]
- Yu D, Zhang J, Wang M, Ji R, Qian H, Xu W. Exosomal miRNAs from neutrophils act as accurate biomarkers for gastric cancer diagnosis. Clin Chim Acta 2024; 554:117773. doi: 10.1016/j.cca.2024.117773 [Crossref] [ Google Scholar]
- Gu J, Xu H, Chen Y, Li N, Hou X. MiR-223 as a regulator and therapeutic target in liver diseases. Front Immunol 2022; 13:860661. doi: 10.3389/fimmu.2022.860661 [Crossref] [ Google Scholar]
- Jeffries J, Zhou W, Hsu AY, Deng Q. miRNA-223 at the crossroads of inflammation and cancer. Cancer Lett 2019; 451:136-41. doi: 10.1016/j.canlet.2019.02.051 [Crossref] [ Google Scholar]
- Wang L, Yang J, Huang J, Wen ZQ, Xu N, Liu X. miRNA expression profile in the N2 phenotype neutrophils of colorectal cancer and screen of putative key miRNAs. Cancer Manag Res 2020; 12:5491-503. doi: 10.2147/cmar.S251427 [Crossref] [ Google Scholar]
- Lee YC, Tzeng WF, Chiou TJ, Chu ST. MicroRNA-138 suppresses neutrophil gelatinase-associated lipocalin expression and inhibits tumorigenicity. PLoS One 2012; 7:e52979. doi: 10.1371/journal.pone.0052979 [Crossref] [ Google Scholar]
- Chen R, Coleborn E, Bhavsar C, Wang Y, Alim L, Wilkinson AN. miR-146a inhibits ovarian tumor growth in vivo via targeting immunosuppressive neutrophils and enhancing CD8 + T cell infiltration. Mol Ther Oncolytics 2023; 31:100725. doi: 10.1016/j.omto.2023.09.001 [Crossref] [ Google Scholar]
- Zhou SL, Yin D, Hu ZQ, Luo CB, Zhou ZJ, Xin HY. A positive feedback loop between cancer stem-like cells and tumor-associated neutrophils controls hepatocellular carcinoma progression. Hepatology 2019; 70:1214-30. doi: 10.1002/hep.30630 [Crossref] [ Google Scholar]
- Ma J, Li N, Lin Y, Gupta C, Jiang F. Circulating neutrophil microRNAs as biomarkers for the detection of lung cancer. Biomark Cancer 2016; 8:1-7. doi: 10.4137/bic.S37333 [Crossref] [ Google Scholar]
- Chen H, Zhang J, He W. The role of microRNAs in γδ T cells: a long way to go. Cell Mol Immunol 2021; 18:2071-2. doi: 10.1038/s41423-021-00711-x [Crossref] [ Google Scholar]
- Sun L, Su Y, Jiao A, Wang X, Zhang B. T cells in health and disease. Signal Transduct Target Ther 2023; 8:235. doi: 10.1038/s41392-023-01471-y [Crossref] [ Google Scholar]
- Chen Z, Zheng Z, Feng L, Huo Z, Huang L, Fu M. Overexpression of miR-382 sensitizes hepatocellular carcinoma cells to γδ T cells by inhibiting the expression of c-FLIP. Mol Ther Oncolytics 2020; 18:467-75. doi: 10.1016/j.omto.2020.07.012 [Crossref] [ Google Scholar]
- Quintavalle C, Incoronato M, Puca L, Acunzo M, Zanca C, Romano G. c-FLIPL enhances anti-apoptotic Akt functions by modulation of Gsk3β activity. Cell Death Differ 2010; 17:1908-16. doi: 10.1038/cdd.2010.65 [Crossref] [ Google Scholar]
- Alkurdi L, Virard F, Vanbervliet B, Weber K, Toscano F, Bonnin M. Release of c-FLIP brake selectively sensitizes human cancer cells to TLR3-mediated apoptosis. Cell Death Dis 2018; 9:874. doi: 10.1038/s41419-018-0850-0 [Crossref] [ Google Scholar]
- Zhu Y, Zhang S, Li Z, Wang H, Li Z, Hu Y. miR-125b-5p and miR-99a-5p downregulate human γδ T-cell activation and cytotoxicity. Cell Mol Immunol 2019; 16:112-25. doi: 10.1038/cmi.2017.164 [Crossref] [ Google Scholar]
- Gordino G, Costa-Pereira S, Corredeira P, Alves P, Costa L, Gomes AQ. MicroRNA-181a restricts human γδ T cell differentiation by targeting Map3k2 and Notch2. EMBO Rep 2022; 23:e52234. doi: 10.15252/embr.202052234 [Crossref] [ Google Scholar]
- Amsen D, Helbig C, Backer RA. Notch in T cell differentiation: all things considered. Trends Immunol 2015; 36:802-14. doi: 10.1016/j.it.2015.10.007 [Crossref] [ Google Scholar]
- Ghorbani S, Talebi F, Chan WF, Masoumi F, Vojgani M, Power C. MicroRNA-181 variants regulate T cell phenotype in the context of autoimmune neuroinflammation. Front Immunol 2017; 8:758. doi: 10.3389/fimmu.2017.00758 [Crossref] [ Google Scholar]
- Gordino GS. Targeting MicroRNAs for Modlation of the Anti-Tumor Human γδ T Cell Differentiation [dissertation]. Universidade de Lisboa; 2016.
- Schmolka N, Papotto PH, Romero PV, Amado T, Enguita FJ, Amorim A. MicroRNA-146a controls functional plasticity in γδ T cells by targeting NOD1. Sci Immunol 2018; 3:eaao1392. doi: 10.1126/sciimmunol.aao1392 [Crossref] [ Google Scholar]
- Hu A, Sun L, Lin H, Liao Y, Yang H, Mao Y. Harnessing innate immune pathways for therapeutic advancement in cancer. Signal Transduct Target Ther 2024; 9:68. doi: 10.1038/s41392-024-01765-9 [Crossref] [ Google Scholar]
- Fiala GJ, Gomes AQ, Silva-Santos B. From thymus to periphery: molecular basis of effector γδ T cell differentiation. Immunol Rev 2020; 298:47-60. doi: 10.1111/imr.12918 [Crossref] [ Google Scholar]
- Haas JD, Nistala K, Petermann F, Saran N, Chennupati V, Schmitz S. Expression of miRNAs miR-133b and miR-206 in the Il17a/f locus is co-regulated with IL-17 production in αβ and γδ T cells. PLoS One 2011; 6:e20171. doi: 10.1371/journal.pone.0020171 [Crossref] [ Google Scholar]
- Ho JY, Hsu RJ, Liu JM, Chen SC, Liao GS, Gao HW. MicroRNA-382-5p aggravates breast cancer progression by regulating the RERG/Ras/ERK signaling axis. Oncotarget 2017; 8:22443-59. doi: 10.18632/oncotarget.12338 [Crossref] [ Google Scholar]
- Bi JG, Zheng JF, Li Q, Bao SY, Yu XF, Xu P. MicroRNA-181a-5p suppresses cell proliferation by targeting Egr1 and inhibiting Egr1/TGF-β/Smad pathway in hepatocellular carcinoma. Int J Biochem Cell Biol 2019; 106:107-16. doi: 10.1016/j.biocel.2018.11.011 [Crossref] [ Google Scholar]
- Zhao J, Nie Y, Wang H, Lin Y. MiR-181a suppresses autophagy and sensitizes gastric cancer cells to cisplatin. Gene 2016; 576:828-33. doi: 10.1016/j.gene.2015.11.013 [Crossref] [ Google Scholar]
- Rezaei T, Hejazi M, Mansoori B, Mohammadi A, Amini M, Mosafer J. microRNA-181a mediates the chemo-sensitivity of glioblastoma to carmustine and regulates cell proliferation, migration, and apoptosis. Eur J Pharmacol 2020; 888:173483. doi: 10.1016/j.ejphar.2020.173483 [Crossref] [ Google Scholar]
- Yao R, Ma YL, Liang W, Li HH, Ma ZJ, Yu X. MicroRNA-155 modulates Treg and Th17 cells differentiation and Th17 cell function by targeting SOCS1. PLoS One 2012; 7:e46082. doi: 10.1371/journal.pone.0046082 [Crossref] [ Google Scholar]
- Hu YZ, Li Q, Wang PF, Li XP, Hu ZL. Multiple functions and regulatory network of miR-150 in B lymphocyte-related diseases. Front Oncol 2023; 13:1140813. doi: 10.3389/fonc.2023.1140813 [Crossref] [ Google Scholar]
- Diener C, Keller A, Meese E. Emerging concepts of miRNA therapeutics: from cells to clinic. Trends Genet 2022; 38:613-26. doi: 10.1016/j.tig.2022.02.006 [Crossref] [ Google Scholar]
- Mardi A, Ghovahi A, Abbasvandi F, Amani D. Experimental validation of miR-4443, miR-572, and miR-150-5p in serum and tissue of breast cancer patients as a potential diagnostic biomarker: a study based on bioinformatics prediction. Biochem Genet 2025. doi: 10.1007/s10528-025-11057-8.
- Rupaimoole R, Wu SY, Pradeep S, Ivan C, Pecot CV, Gharpure KM. Hypoxia-mediated downregulation of miRNA biogenesis promotes tumour progression. Nat Commun 2014; 5:5202. doi: 10.1038/ncomms6202 [Crossref] [ Google Scholar]
- Shah MY, Calin GA. MicroRNAs as therapeutic targets in human cancers. Wiley Interdiscip Rev RNA 2014; 5:537-48. doi: 10.1002/wrna.1229 [Crossref] [ Google Scholar]
- Ho PT, Clark IM, Le LT. MicroRNA-based diagnosis and therapy. Int J Mol Sci 2022; 23:7167. doi: 10.3390/ijms23137167 [Crossref] [ Google Scholar]
- Shah MY, Ferrajoli A, Sood AK, Lopez-Berestein G, Calin GA. microRNA therapeutics in cancer - an emerging concept. EBioMedicine 2016; 12:34-42. doi: 10.1016/j.ebiom.2016.09.017 [Crossref] [ Google Scholar]
- Zhang X, Li Y, Chen YE, Chen J, Ma PX. Cell-free 3D scaffold with two-stage delivery of miRNA-26a to regenerate critical-sized bone defects. Nat Commun 2016; 7:10376. doi: 10.1038/ncomms10376 [Crossref] [ Google Scholar]
- Katoh S, Yoshioka H, Senthilkumar R, Preethy S, Abraham SJK. Enhanced miRNA-140 expression of osteoarthritis-affected human chondrocytes cultured in a polymer based three-dimensional (3D) matrix. Life Sci 2021; 278:119553. doi: 10.1016/j.lfs.2021.119553 [Crossref] [ Google Scholar]
- Samad AF, Kamaroddin MF, Sajad M. Cross-kingdom regulation by plant microRNAs provides novel insight into gene regulation. Adv Nutr 2021; 12:197-211. doi: 10.1093/advances/nmaa095 [Crossref] [ Google Scholar]
- Wang P, Zhou Y, Richards AM. Effective tools for RNA-derived therapeutics: siRNA interference or miRNA mimicry. Theranostics 2021; 11:8771-96. doi: 10.7150/thno.62642 [Crossref] [ Google Scholar]
- Montgomery RL, Yu G, Latimer PA, Stack C, Robinson K, Dalby CM. MicroRNA mimicry blocks pulmonary fibrosis. EMBO Mol Med 2014; 6:1347-56. doi: 10.15252/emmm.201303604 [Crossref] [ Google Scholar]
- Yuen JG, Hwang GR, Fesler A, Intriago E, Pal A, Ojha A. Development of gemcitabine-modified miRNA mimics as cancer therapeutics for pancreatic ductal adenocarcinoma. Mol Ther Oncol 2024; 32:200769. doi: 10.1016/j.omton.2024.200769 [Crossref] [ Google Scholar]
- Hong DS, Kang YK, Borad M, Sachdev J, Ejadi S, Lim HY. Phase 1 study of MRX34, a liposomal miR-34a mimic, in patients with advanced solid tumours. Br J Cancer 2020; 122:1630-7. doi: 10.1038/s41416-020-0802-1 [Crossref] [ Google Scholar]
- Beg MS, Brenner AJ, Sachdev J, Borad M, Kang YK, Stoudemire J. Phase I study of MRX34, a liposomal miR-34a mimic, administered twice weekly in patients with advanced solid tumors. Invest New Drugs 2017; 35:180-8. doi: 10.1007/s10637-016-0407-y [Crossref] [ Google Scholar]
- Bouchie A. First microRNA mimic enters clinic. Nat Biotechnol 2013; 31:577. doi: 10.1038/nbt0713-577 [Crossref] [ Google Scholar]
- Daige CL, Wiggins JF, Priddy L, Nelligan-Davis T, Zhao J, Brown D. Systemic delivery of a miR34a mimic as a potential therapeutic for liver cancer. Mol Cancer Ther 2014; 13:2352-60. doi: 10.1158/1535-7163.Mct-14-0209 [Crossref] [ Google Scholar]
- Kelnar K, Bader AG. A qRT-PCR method for determining the biodistribution profile of a miR-34a mimic. Methods Mol Biol 2015; 1317:125-33. doi: 10.1007/978-1-4939-2727-2_8 [Crossref] [ Google Scholar]
- van der Ree MH, de Vree JM, Stelma F, Willemse S, van der Valk M, Rietdijk S. Safety, tolerability, and antiviral effect of RG-101 in patients with chronic hepatitis C: a phase 1B, double-blind, randomised controlled trial. Lancet 2017; 389:709-17. doi: 10.1016/s0140-6736(16)31715-9 [Crossref] [ Google Scholar]