Bioimpacts. 16:32564.
doi: 10.34172/bi.32564
Review
The interplay between non-coding RNAs and Wnt signaling pathway in leukemia: A narrative review
Reza Biglari Conceptualization, Writing – original draft, 1, 2 
Majid Farshdousti Hagh Conceptualization, Funding acquisition, 2
Khadijeh Abbasi Conceptualization, Visualization, 1
Maghsood Shaaker Conceptualization, Investigation, 1
Amir Mehdizadeh Conceptualization, Methodology, Supervision, Validation, Writing – review & editing, 2, *
Author information:
1Department of Biochemistry and Clinical Laboratories, Faculty of Medicine, Tabriz University of Medical Sciences, Tabriz, Iran
2Hematology and Oncology Research Center, Tabriz University of Medical Sciences, Tabriz, Iran
Abstract
Leukemias are heterogeneous hematologic malignancies that involve dysregulated proliferation and impaired differentiation of hematopoietic cells. The Wnt signaling pathway, which regulates hematopoietic stem cell self-renewal and lineage commitment, is dysregulated across leukemia subtypes. Non-coding RNAs (ncRNAs), which include microRNAs, long non-coding RNAs (lncRNAs), and circular RNAs, have been identified as important post-transcriptional and epigenetic regulators of oncogenic signaling networks. There is growing evidence for a complex, bidirectional relationship between ncRNAs and the Wnt/β-catenin pathway, but this has not yet been summarized or integrated into the context of leukemia. This study examines how ncRNA manipulation of core Wnt components – β-catenin, Frizzled receptors, and Dvl (dishevelled) proteins – affect leukemic cell survival, proliferation, stemness, and treatment resistance. Furthermore, we discuss reciprocal regulation, in which Wnt stimulation affects ncRNA production, forming feed-forward loops that promote leukemogenesis. By synthesizing disparate findings from studies investigating ncRNA and Wnt signaling mechanisms across leukemia subtypes, we identify critical mechanistic gaps in the literature, as well as opportunities and controversies that could be that could be leveraged in evaluating ncRNA–Wnt interactions as diagnostic and therapeutic targets. This review presents a comprehensive study integrating ncRNA biology and Wnt-driven leukemic development in order to identify crucial insights into disease vulnerabilities and future research initiatives.
Graphical Abstract
Keywords: Leukemia, Wnt signaling, Non-coding RNAs, Drug resistance
Copyright and License Information
© 2026 The Author(s).
This work is published by BioImpacts as an open access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (
http://creativecommons.org/licenses/by-nc/4.0/). Non-commercial uses of the work are permitted, provided the original work is properly cited.
Funding Statement
This study was supported by Hematology and Oncology Research Center at Tabriz University of Medical Sciences, Tabriz, Iran (Grant No.: 71742).
Introduction
Leukemia pathogenesis begins when somatic genetic and epigenetic alterations accumulate in hematopoietic stem or progenitor cells, leading to unrestricted proliferation and abnormal differentiation.1,2 These molecular alterations disrupt normal cellular processes, including growth, survival, and apoptosis, which leads to an expansion of transformed clonal hematopoietic cells that infiltrate the bone marrow and peripheral blood.1,3 Common leukemogenic drivers include recurrent chromosomal abnormalities, such as translocations observed in chronic myeloid leukemia (CML) and acute lymphoblastic leukemia (ALL) that generate oncogenic fusion genes.4,5 Although there are recurrent mutations that create oncogenic fusion genes, some of the most common mutations occur in inherited mutations, transcription factors, genes involved in relevant signaling pathways (i.e., FLT3, RAS), or epigenetic modifications that are referred to as mutational events (i.e., mutations to DNMT3A, TET2). These and other mutations likely contribute to the varied genomic diversity of acute myeloid leukemia (AML) and other myeloid neoplasms.2,3,6 The proliferative and survival advantages of the leukemic cells is derived in part from a dysregulated bone marrow microenvironment that promotes a supportive niche, that enables leukemic cell proliferation and drug resistance.1,2 Understanding the interplay between intrinsic genetic alterations and the extrinsic environmental factors is needed to clarify the pathways within the clonal expansion of leukemic transformation and to develop a targeted therapeutic approach to treating the diverse threats presented by chronic and acute leukemias.1,2,6
The Wnt signaling pathway, which is a well-conserved pathway that is essential for normal hematopoiesis and embryonic development, plays a complicated role in the development of leukemia. Aberrant activation of Wnt signaling due to loss of regulation is one of the most common events in various types of leukemia. Activation of the Wnt pathway can promote the self-renewal, survival, and proliferation of leukemic cells.7,8 Canonical Wnt/β-catenin activation is thought to enhance LSC maintenance, which has important implications with disease initiation, progression, and relapse particularly in acute myeloid leukemia.8,9 Wnt signaling may also modulate the bone marrow microenvironment creating a niche that supports leukemic cell survival or proliferation, thereby enabling persistence and resistance. 7 Beyond the potential direct effects of Wnt signaling in leukemic cells, little is known about how it contributes to the complex roles the Wnt signaling in stem cell biology and acute leukemia development. Nevertheless, targeting this pathway may open new avenues for developing treatments for this challenging disease.8,9
ncRNAs have been shown to be significant regulatory factors in the Wnt signaling pathways in leukemia, contributing to an intricate network of factors that profoundly influence disease development
ncRNAs have been shown to be significant regulatory factors in the Wnt signaling pathways in leukemia, contributing to an intricate network of factors that are profoundly influence disease development.
miRNAs, a class of ncRNAs, are most often considered in targeting Wnt components. For example, in chronic lymphocytic leukemia (CLL), miRNAs have been shown to target intrinsic regulators of WNT signaling, and in CML, the expression of sFRP1 can promote miRNAs that target Wnt signaling.10,11
Similarly, long non-coding RNAs play roles in CML, with lncRNA SNHG1 promoting acute myeloid leukemia cell proliferation by regulating the miR-489-3p/SOX12/Wnt/β-catenin axis.12 Certain circRNAs, such as hsa_circ_0121582, can suppress leukemia development by antagonizing Wnt/β-catenin signaling.13 Other complex interactions have emerged, demonstrating that ADAR1's activity is critical for acute myeloid leukemia cell survival. It achieves this by inhibiting miRNA biogenesis, which regulates post-transcriptional WNT signaling.14 Understanding these reciprocal regulatory relationships between ncRNAs and Wnt signaling offers valuable opportunities to identify new therapeutic targets for leukemia. This narrative review will provide a thorough synthesis of the current understanding of the complex relationship between ncRNAs and the Wnt signaling pathway in leukemia. We will address how ncRNAs regulate Wnt signaling and vice versa, as well as the impact of Wnt activation on ncRNA expression. A better understanding of the regulatory complexity of ncRNAs and the Wnt signaling pathway provides greater insight into leukemia pathogenesis of and aids in the identification of novel diagnostic biomarkers and therapeutic targets.
The Wnt signaling pathway in leukemia: An overview
Canonical Wnt/β-catenin pathway
The canonical Wnt/β-catenin pathway is a central signaling pathway regulating important functions during embryonic development, tissue homeostasis, and stem cell maintenance, including hematopoietic stem cells.15,16 Careful regulation of the pathway is essential, as its inappropriate activation is linked to various diseases, particularly hematological malignancies such as leukemia.15,17 The pathway begins with the binding of secreted Wnt ligands to transmembrane Frizzled (FZD) receptors and low-density lipoprotein receptor-related protein (LRP5/6) co-receptors expressed on the cell surface. This binding activates downstream signaling pathways that subsequently result in inhibition of a large cytoplasmic "destruction complex" composed of Axin, adenomatous polyposis coli (APC), glycogen synthase kinase 3 beta (GSK3β), and casein kinase 1 alpha (CK1α). In the absence of stimulation with Wnt ligands, this destruction complex phosphorylates β-catenin, tagging it for ubiquitination and degradation by the proteasome, keeping cytoplasmic β-catenin levels low. Once cells signal through the Wnt pathway, the destruction complex is disassembled or inactivated, and β-catenin is allowed to accumulate in the cytoplasm. Once stabilized, catenin enters the nucleus, where it binds to TCF/LEF transcription factors. This interaction transforms TCF/LEF from transcriptional repressors into activators, inducing the expression of many Wnt target genes that are involved in cell proliferation, survival, differentiation, and self-renewal, including c-MYC and CCND1 (Cyclin D1).15
In hematologic neoplasms, particularly leukemia, the dysregulation of the Wnt/β-catenin pathway is an important oncogenic event.15,17 This aberrant activation helps sustain both uncontrolled proliferation and survival of leukemic cells and plays an important role in sustaining the self-renewal potential of leukemia stem cells (LSCs), which promote initiation, progression, and relapse of these diseases.15,17 For example, in AML, the Wnt/β-catenin pathway is often constitutively active and contributes to the aggressive nature of the disease and therapy resistance, as this pathway often acts downstream of enhancer regions in regularly activated signaling pathways. This hyperactivation can be the result of many potential mechanisms, including mutations in various components of the pathway (e.g., APC, AXIN1, CTNNB1 (β-catenin)) and/or the overexpression of Wnt ligands or receptors.15 In addition, the stability and activity of β-catenin are more than simply transcriptional. It can interact with conventional RNA-binding proteins (RBPs), including Musashi-2 (MSI2), to co-occupy a Wnt signaling mRNA network in myeloid leukemia cells. This network regulates important post-transcriptional gene programs required for leukemogenesis.18
Such a broad regulatory role suggests that β-catenin regulates not only transcription but also mRNA processing and translation, reinforcing our view of it as a central player in myeloid leukemias. Similarly, in T-cell acute lymphoblastic leukemia (T-ALL), the Wnt/β-catenin pathway is often hyperactivated, sometimes alongside other oncogenic pathways such as PI3K/Akt/mTOR, leading to dysregulated cell proliferation and survival.17
Overall, the Wnt/β-catenin pathway represents a promising therapeutic target in leukemia due to its central role in promoting leukemic cell proliferation and maintaining stem cell identity.17,19 Potential agents targeting the Wnt/β-catenin pathway are under development, preclinical studies are beginning to show promising results. For instance, the multi-CDK inhibitor dinaciclib has been shown to reverse resistance to bromo- and extra-terminal domain (BET) inhibitors in acute myeloid leukemia by inhibiting Wnt/β-catenin signaling. This suggests that combining dinaciclib with other agents could enhance therapeutic efficacy against AML.19 Understanding of the complex mechanisms by which Wnt signaling becomes dysregulated and contributes to leukemic transformation continues to grow. Further discoveries of its interactions with cellular processes and critical regulators, including RBPs, may enable the development of targeted therapies for these challenging hematological malignancies.15,18
Non-canonical Wnt pathways
The non-canonical Wnt signaling pathway, which includes the distinct Wnt/PCP (planar cell polarity) and Wnt/β-catenin pathways, is increasingly recognized as important in the development of leukemias and other hematological malignancies. Although the evidence is rapidly emerging, the distinct β-catenin-independent cascades may provide therapies typically targe Wnt signaling when altered in the context of hematological malignancies.
Within the context of adult T-cell leukemia/lymphoma (ATL), a very aggressive malignancy caused by human T-cell leukemia virus type 1 (HTLV-1) infection, Wnt5a is aberrantly overexpressed contributing to the malignant phenotype.20 Wnt5a, as an important ligand involved in non-canonical Wnt signaling, has been shown to enhance ATL cell proliferation and survival, and possibly prevent apoptosis.20 These reports imply, targeting the Wnt5a-mediated non-canonical cascade may be an avenue for significantly reducing or blocking oncogenesis by controlling major malignant attributes associated with ATL.20
Additional information pertaining to non-canonical pathway regulation in leukemia stems from studies in CML. For example, Dvl (dishevelled) proteins are important cytoplasmic scaffold proteins that differentially regulate canonical and non-canonical Wnt signaling pathways. 21 Studies have also shown that Dvl proteins modulate the expression of several other important proteins (e.g., in CML cell lines, we have previously identified pathways for Dvl, including SMAD1, Aryl Hydrocarbon Receptor (AHR), mammalian target of rapamycin (mTOR), and Bromodomain-containing protein 7 (BRD7)). 21 Such studies suggest a complex relationship where Dvl-mediated non-canonical Wnt signaling might functionally crosstalk with other survival and proliferation-relevant signaling pathways, such as mTOR, a central regulator of cell growth and metabolism.21 Dvl proteins allow integration of several distinct targets and pathways, reflecting the complex nature of the regulatory network controlled by non-canonical Wnt signaling and implications for therapeutic targeting of CML.21
Together, these findings show the importance of non-canonical Wnt signaling in leukemia. The potential contributions of specific ligands, like Wnt5a, to ATL and Dvl proteins, indicate the contribution of this pathway in disease progression. Targeting these aberrant mechanisms of non-canonical Wnt signaling, either by inhibition of Wnt5a and its signaling or by reducing the contribution of Dvl proteins, represents a potential avenue for new targeted therapies for various leukemias. Fig. 1 illustrates the canonical and non-canonical Wnt signaling dysregulation in leukemias.
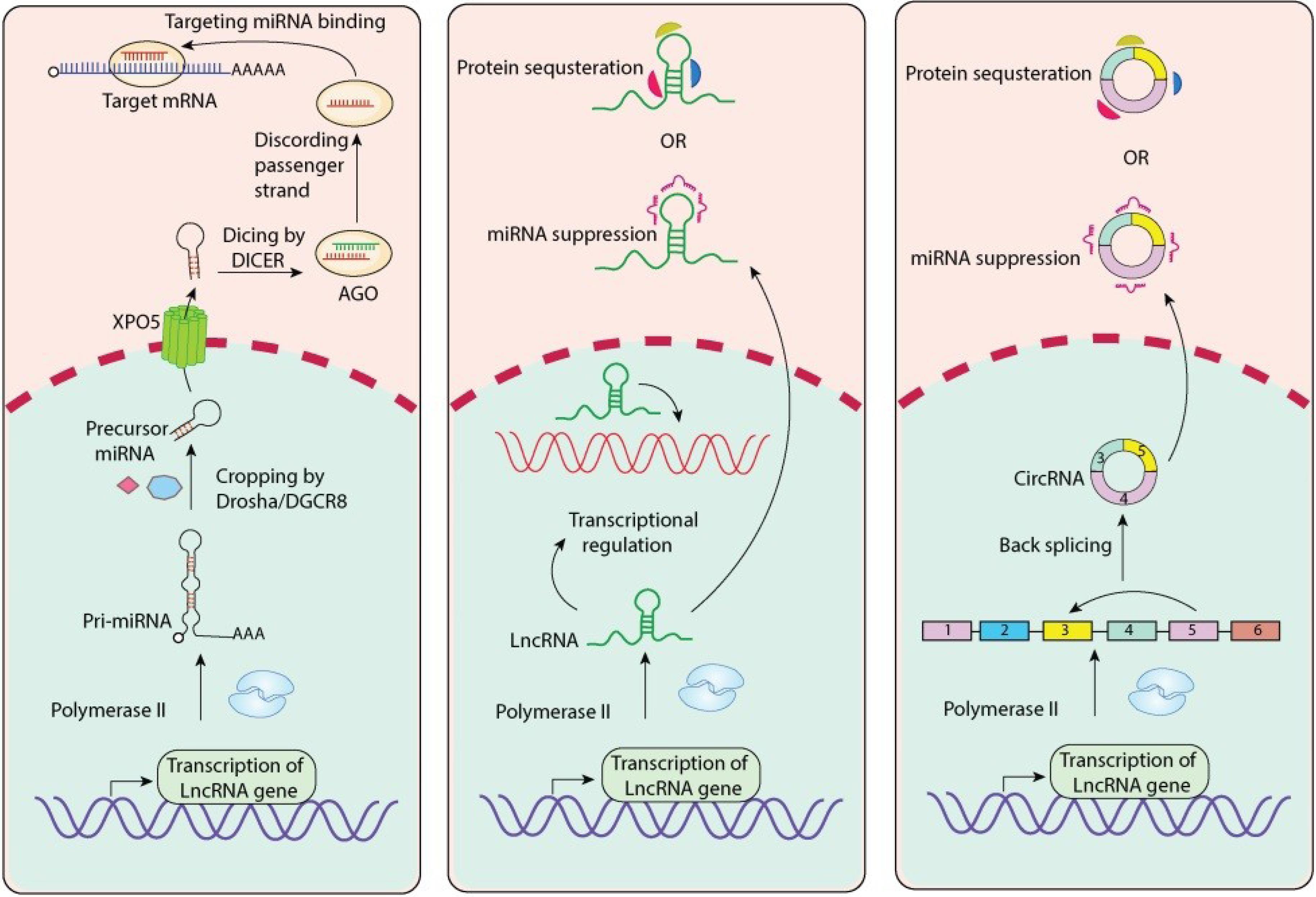
Fig. 1.
Overview of Canonical and Non-Canonical Wnt Signaling Pathways and Their Therapeutic Targets. The Wnt signaling pathway splits into two main branches, continuing with the canonical (β-catenin-dependent) pathway (left) and the non-canonical (β-catenin-independent) pathway (right). For the canonical pathway, Wnt ligands bind to Fzd receptors and LRP5/6 co-receptors, subsequently recruiting dishevelled (Dvl) and inhibiting the β-catenin destruction complex consisting of APC, Axin, CK1, and GSK3β. The inhibited β-catenin is stabilized, translocates into the nucleus, and activates gene transcription in complex with LEF1/TCF. Several upstream target inhibitors of the canonical Wnt signaling pathway are presented, including BHQ880, DKN-01, and DKKs, which all inhibit the interaction between Wnt-LRP, BC2059, and CWP232291. They take a different approach and interrupt β-catenin instead; PRI-724 is able to interrupt β-catenin signaling without blocking its interaction. The non-canonical pathway works in a β-catenin-independent manner and uses different co-receptors, for example, ROR1/2. Binding of Wnt activates the downstream effectors through Dishevelled and activates the Wnt/Ca2+ pathway (through PLC, PKC, and NFAT) and the planar cell polarity (PCP) pathway (through Daam1, RhoA, Rock, and JNK), ultimately affecting cytoskeletal dynamics and gene expression. Cirmtuzumab is an anti-ROR1 therapeutic antibody directed against this non-canonical pathway. The remaining extracellular regulators are sFRPs and WIF that sequester Wnt ligands, Norrin (a different ligand for Fzd), RSPO (potentiates Wnt signaling through LGR4/5), and ZNFR3 (a negative regulator of receptor availability). Therapeutic inhibitors are represented with the pill icons, as are some components of specific pathways, and we've used colors for clarity. The figure was generated by Adobe Illustrator 30.0.0.123 software.
.
Overview of Canonical and Non-Canonical Wnt Signaling Pathways and Their Therapeutic Targets. The Wnt signaling pathway splits into two main branches, continuing with the canonical (β-catenin-dependent) pathway (left) and the non-canonical (β-catenin-independent) pathway (right). For the canonical pathway, Wnt ligands bind to Fzd receptors and LRP5/6 co-receptors, subsequently recruiting dishevelled (Dvl) and inhibiting the β-catenin destruction complex consisting of APC, Axin, CK1, and GSK3β. The inhibited β-catenin is stabilized, translocates into the nucleus, and activates gene transcription in complex with LEF1/TCF. Several upstream target inhibitors of the canonical Wnt signaling pathway are presented, including BHQ880, DKN-01, and DKKs, which all inhibit the interaction between Wnt-LRP, BC2059, and CWP232291. They take a different approach and interrupt β-catenin instead; PRI-724 is able to interrupt β-catenin signaling without blocking its interaction. The non-canonical pathway works in a β-catenin-independent manner and uses different co-receptors, for example, ROR1/2. Binding of Wnt activates the downstream effectors through Dishevelled and activates the Wnt/Ca2+ pathway (through PLC, PKC, and NFAT) and the planar cell polarity (PCP) pathway (through Daam1, RhoA, Rock, and JNK), ultimately affecting cytoskeletal dynamics and gene expression. Cirmtuzumab is an anti-ROR1 therapeutic antibody directed against this non-canonical pathway. The remaining extracellular regulators are sFRPs and WIF that sequester Wnt ligands, Norrin (a different ligand for Fzd), RSPO (potentiates Wnt signaling through LGR4/5), and ZNFR3 (a negative regulator of receptor availability). Therapeutic inhibitors are represented with the pill icons, as are some components of specific pathways, and we've used colors for clarity. The figure was generated by Adobe Illustrator 30.0.0.123 software.
ncRNAs in leukemia: Classification and roles
The molecular biology landscape in cancer has undergone a paradigm shift with the discovery of ncRNAs being key modulators of transcription and cellular activities. The ncRNAs were long regarded as transcriptional noise, and are, in fact, not protein-coding. Until recently, the roles of ncRNAs in health and disease were largely misunderstood, but they have now been shown to have diverse and significant roles, including the initiation, progression, and response to therapy of a multitude of malignancies. Leukemia, a complex and heterogeneous group of hematological malignancies characterized by excessive and uncontrolled proliferation of aberrant blood cells, represents a robust area where ncRNAs play significant roles. 22 Dysregulation of ncRNAs is directly associated with deregulation of transcriptional and other key oncogenic signatures that affect on programs regulating cellular proliferation, differentiation, apoptosis, and the immune microenvironment.22,23 The present section will first classify the major types of ncRNAs associated with leukemias and provide a brief overview of their roles, which may be characterized and utilized as diagnostic and prognostic biomarkers and new therapeutic targets.
Classification of ncRNAs in leukemia
ncRNAs can be classified into broad categories depending on their size and structure. The most studied non-coding RNAs in the setting of leukemia are miRNAs, lncRNAs, and circRNAs.
miRNAs
MicroRNAs are a class of short (generally 19-25 nucleotides), evolutionarily conserved ncRNAs, and they post-transcriptionally regulate gene expression.24 miRNAs execute gene regulation by base pairing with the complementary sequence, mostly in the 3' untranslated regions (3'UTRs) of their target messenger RNAs (mRNAs), which ultimately leads to either translational repression or mRNA destruction.24 miRNA biogenesis involves multiple enzymes from the generation of miRNA transcripts to the formulation of mature and functional miRNAs, including Drosha and Dicer, for example.
MiRNAs are often dysregulated in leukemia, either as onco-miRs or as tumor suppressors.24,25 For instance, the miR-15a/miR-16-1 cluster, which is located in the DLEU2 locus, is often deleted or downregulated in CLL, which accounts for uncontrolled cell proliferation and apoptosis suppression, thus demonstrating their tumor-suppressive role.25,26 undoubtedly, some other miRNAs are found to be aberrantly upregulated in leukemia and would promote leukemogenesis. An example of this is miR-125b, which has been observed to promote and accelerate PML-RARα-mediated murine acute promyelocytic leukemia (APL),27 which is a subtype of AML. MicroRNA-22 (miR-22) has been shown to have a complex role in leukemia, with involvement in apoptosis, proliferation, and therapeutic response, and has even been assessed for prognostic significance in leukemia patients.28,29 In AML, miR-185-5p has demonstrated the ability to inhibit leukemic cell growth by suppressing GPX1.30 Additionally, engineered mesenchymal stem cell exosomes containing miR-34c-5p have been successful in selectively facilitating the destruction of acute myeloid leukemia stem cells (LSCs), establishing the potential therapeutic implications of miRNAs.31 In the case of acute ALL, miR-126 is associated with a subset of quiescent and chemoresistant human B-ALL cells that correspond with minimal residual disease,32 and the inhibition of miR-126 has been implemented in specific Ph + ALL studies.33
lncRNAs
LncRNAs are ncRNA molecules longer than 200 nucleotides and are not translated into proteins. Although lncRNAs are not protein-coding molecules, they are structurally diverse and exhibit functional specificity through their effects on gene regulation, including chromatin remodeling, transcriptional regulation, post-transcriptional modification, and competing endogenous RNAs (ceRNAs).23 Unlike miRNAs and other ncRNAs, the functional mechanisms of lncRNAs are highly context dependent (e.g., act as scaffolds that recruit protein complexes to specific genomic regions, serving as decoys that absorb transcription factors, or absorbing miRNAs or their targets).23,34
LncRNAs continue to show their potential to be relevant to the pathogenesis, progression, and drug resistance in leukemia.23,34,35 A common characteristic among diverse subtypes of leukemia is the dysregulation of lncRNAs.36,37 For example, some lncRNAs in stem cells are known to be involved in the development of leukemia, suggesting their role in maintaining the malignant stem cell phenotype.34 LncRNAs can modulate treatment outcomes in leukemia, with evidence in studies of their involvement in drug resistance mechanisms.38,39 In the case of AML, some lncRNAs, such as LINC00987 (lncRNA), have been shown to reverse adriamycin drug resistance by changing the regulation of the miR-4458/HMGA2 axis.40 Other lncRNAs, such as XIST, have been reported to affect AML cell function via the miR-142-5p-PFKP axis.41 The lncRNA HOTAIRM1 (HOX antisense intergenic RNA myeloid 1) from the HOXA cluster regulates myeloid differentiation; the dysregulation of this lncRNA is involved in AML.42 LncRNAs in ALL have also been shown to regulate cellular traits essential for leukemogenesis.43 The bone marrow osteoblast mesenchymal stromal cell exosomes have recently also been studied as a component of the bone marrow microenvironment, with reports on biologically relevant lncRNAs transferring via exosomes to regulate myeloid cells.44
circRNAs
Distinct from linear versions, circular RNAs are a new type of ncRNA with covalently closed loop structures that do not have 5' caps or 3' poly(A) tails, and hence they are connected to unique aspects of mRNA metabolism and gene regulation.45 Circular RNAs’ unique structure and closure make them particularly resilient to degradation by exonucleases.45 CircRNAs are believed to be generated by backsplicing when a downstream 5' splice site is joined with an upstream 3' splice site.46
Early research has shown that circRNAs likely play a significant role in the development of leukemia and leukemia progression.45,47,48 The primary function of circRNA is often thought to be "miRNA sponges," where circRNAs sequester miRNAs with the effect of derepressing miRNA target genes.45,46 Circular RNAs have been extensively studied in AML; it was shown that circRNAs can be interplay with oncogenic and tumor-suppressive functions by sponging miRNAs that can be oncogenic or tumor-suppressive.46,49
CircPVT1, for example, has been described to support AML malignancy, with MEF2A as its transcription factor. Although circRNAs and untranslatable regions (UTRs) are emerging in our understanding of acute myeloid leukemia, they've gained attention in the context of disease progression.49 The circRNAome of childhood ALL has been defined, providing subtype-specific expression profiles and indicating involvement in specific fusion-driven leukemias such as TCF3/PBX1 ALL.50 circRNAs can also bind directly to RNA-binding proteins, act as scaffolds for proteins, or even be translated into small peptides in certain contexts, but most of the research in leukemia has focused on circRNA activities as ceRNAs as compared to other interactions with cellular components.45,46 The concept of ceRNAs, which include lncRNAs and circRNAs, builds and integrate an even more complex regulatory network in AML that has biologically and clinically relevant implications.48 Table 1 summarizes the ncRNAs in leukemia biology.
Table 1.
The role of ncRNAs in leukemias
|
ncRNA
|
Leukemia type
|
Function/Mechanism
|
Clinical relevance
|
Ref.
|
| uc.112 |
Childhood T/B-ALL |
Overexpressed in T-ALL vs B-ALL |
Potential diagnostic marker |
51
|
| SNHG family |
AML |
Multiple members associated with prognosis |
Prognostic biomarkers |
52
|
| GAS5 |
AML |
Polymorphism impacts hematopoietic recovery |
Poor prognosis in Chinese AML patients |
53
|
| MALAT1 |
AML, CLL |
Promotes proliferation, migration, suppresses apoptosis via miR-146a/CXCR4 |
Poor prognosis |
12,54,55
|
| ZEB2-AS1 |
AML |
Sponges miR-122-5p → activates PLK1 pathway |
Promotes proliferation, inhibits apoptosis |
56
|
| TUG1 |
AML |
Upregulation promotes proliferation via AURKA |
Correlates with poor prognosis |
57-59
|
| HOXA-AS2 |
AML |
| Identified through RNA-seq, correlated with clinical outcomes |
Potential prognostic biomarker |
60
|
| HOXB-AS3 |
AML, MDS |
| Promotes proliferation |
Adverse prognostic factor |
61-63
|
| LINC00152 |
AML |
Regulates leukemia stem cell self-renewal and chemo-resistance |
Therapeutic target |
64
|
| LINC00641 |
AML |
miR-378a/ZBTB20 axis |
Enhances proliferation and migration |
65
|
| LINC00662 |
AML |
Sponges miR-340-5p to activate ROCK1 |
Contributes to malignancy |
66
|
| LINC01268 |
AML |
Promotes growth, suppresses apoptosis via miR-217/SOS1 |
Prognostic marker |
67,68
|
| LINC00899 |
AML |
Detected in serum |
Novel biomarker |
69
|
| LINC00987 |
AML |
Sponges miR-4458 → regulates HMGA2 |
Overcoming drug resistance |
40
|
| UCA1 |
AML, APL |
Induced by C/EBPα-p30, sponges miR-126 |
Promotes proliferation, diagnostic marker |
70,71
|
| NEAT1 |
ALL |
Sponges miR-335-3p |
Related to poor prognosis |
55
|
| CCAT1 |
AML |
Acts as ceRNA |
| Regulates growth and differentiation |
72
|
| HOTAIR |
AML |
Demethylates HOXA5 via Dnmt3b inhibition |
Tumor suppressor function when silenced |
73
|
| KIAA0125 |
AML |
High expression profile |
Distinct clinical characteristics |
74
|
| CASC15 |
AML |
Induced by IDH mutations, maintains immature phenotype |
Pathogenic role |
75
|
| IRAIN |
CML |
Inhibits NF-κB/CD44 → Overcomes imatinib resistance |
Therapeutic relevance |
76
|
| BLACE |
B-ALL |
Diagnostic ncRNA |
Biomarker candidate |
77
|
| RP11-252C15.1 |
B-ALL |
Hallmark of leukemogenesis in children |
Prognostic marker |
78
|
| AC002454.1 |
Pediatric AL |
Clinical relevance under investigation |
Diagnostic/prognostic potential |
79
|
| MBNL1-AS1 |
AML |
High expression correlates with prognosis |
Prognostic value |
80
|
| DLEU2/DLEU2L |
CLL, AML, Pancreatic Cancer |
Tumor suppressor, modulates chemo-resistance |
Epigenetically silenced in CLL |
81-83
|
| IRF2-3 / KIAA1755-4 |
CLL |
High expression = poor prognosis |
Prognostic indicators |
84
|
| Lnc-223 |
AML |
Host of miR-223, acts as ceRNA to induce IRF4 |
Regulatory role in AML development |
85
|
| KCNQ1OT1 |
AML |
Retracted due to data issues |
N/A |
86,87
|
| CASC15 |
AML |
Associated with IDH mutations |
Maintains stem-like state |
75
|
| circRNAs |
AML, CLL, BM microenvironment |
Regulate gene expression, epigenetics |
Potential therapeutic targets |
88-90
|
| Various miRNAs (e.g., miR-335-3p, miR-126) |
AML, ALL, CML |
Modulated by lncRNAs such as MALAT1, NEAT1, UCA1 |
Drug resistance, survival, differentiation |
55,91-93
|
The interplay: ncRNAs as modulators of Wnt signaling in leukemia
The Wnt/β-catenin signaling pathway is a major regulator of cellular proliferation, differentiation, and survival. In hematological malignancies, such as leukemia, dysregulation of this pathway contributes to disease initiation, progression, and therapy resistance. Recently, a growing body of evidence has established that ncRNAs, such as miRNAs, lncRNAs, and circRNAs, can play a role in regulating Wnt signaling and thereby contribute to leukemogenesis. In this section, we review the emerging literature describing how ncRNAs regulate the Wnt/β-catenin axis in various forms of leukemia, including mechanistic insights and potential therapeutic directions.
miRNAs as modulators of Wnt signaling in leukemia
Although most studies characterize miRNAs as regulators of Wnt/β-catenin signaling in leukemia, limitations remain in the reported outcomes, mechanisms of action, and context-specific finding. For example, miR-155 is frequently reported as an oncogenic activator of Wnt signaling in ALL and AML drug resistance.76,94 However, it is established in other contexts that miR-155 has immunomodulatory, or even tumor-suppressive, activities. Furthermore, these studies do not usually address how the leukemia lineage, pro-inflammatory signals, and/or mutational background may influence the miR-155 effects on Wnt signaling, leaving many questions about the functional role of miR-155. Claims regarding miR-370-3p95 and miR-222-3p96 describe them as an inhibitor and an agonist of Wnt signaling, respectively, yet still leave the question of whether Wnt modulation is actually necessary or sufficient to drive these anti-leukemic or pro-leukemic effects. In most of these studies, involvement of the Wnt pathway was inferred only from changes in β-catenin expression and/or Wnt reporter activity, while functional rescue of Wnt pathway activity following miRNA perturbation has been demonstrated only rarely.
A further key limitation is the recurring assumption that miRNAs act via single targets (like USP48 for miR-26b-5p97 or SOX15 for miR-129498). Because of miRNAs’ pleiotropic tendency, such models are almost certainly incomplete and do not necessarily address the resulting contribution of Wnt signaling to phenotype generation and treatment responses. For example, with the miR-99a/100~125b tricistron,99 which modestly seems to balance TGFβ–Wnt signaling, additional broad transcriptional, metabolic, and epigenetic consequences are not explicitly considered. Most data, if not all, are generated from leukemia cell lines and lack widespread verification in primary patient samples or in in vivo studies, and whether miRNA–Wnt associations and regulatory mechanisms persist in the varying structural or biological conditions of leukemia remains unclear. Furthermore, it is currently unclear if “miRNA–Wnt signatures” exist across leukemia lineages or states of resistance, or if they can be identified as stable therapeutic or diagnostic markers.
lncRNAs: Multifunctional regulators of the Wnt pathway
Studies looking at lncRNAs have led to a similarly complex and inconsistent picture around their modulation of Wnt signaling. Most studies examined lncRNAs such as SNHG1,100 LINC00909,101 and DUXAP8102 through ceRNA-based models, wherein they were assumed to predict modulation of Wnt activity by sequestering a single miRNA, but to generate a functional ceRNA model for an lncRNA, the functional abundance of the lncRNA compared to its targeted miRNA must be known. When examining the efficiency of ceRNA models, relative abundance is often estimated based on qPCR testing of patient samples, rather than measuring the abundances of lncRNA and competing miRNAs directly in patient-derived leukemia cells. Therefore, the biological plausibility of ceRNA-mediated Wnt modulation is unclear in most of the cases reported. In addition, other potential lncRNA functions, including chromatin remodeling, scaffolding functions, or direct interactions with Wnt signaling proteins, are rarely accounted for in many of the studies reported, even though lncRNA scaffolding effects and roles in chromatin regulation have clearly been demonstrated in other cancer types.
These methodological limitations are particularly concerning when interpreting both metabolic and Wnt interactions mediated by lncRNA. For example, DUXAP8102 is said to inhibit glycolysis due to Wnt inhibition, while HOTAIRM1 appears to promote glycolytic flux as a result of Wnt activation in AML.75,103 Together, these observations suggest that there is no universally applicable mechanistic model connecting Wnt signaling to metabolic modulation in leukemia. This raises the possibility that other microenvironmental or genetic determinants specify how Wnt is integrated into metabolic processes. LncRNAs such as HOTTIP104 and LEF-AS1105 have been identified as regulators of hematopoietic progenitor function, but their direct involvement in Wnt signaling is not well supported. Therefore, their involvement in Wnt signaling may allow for alternative explanations, such as regulating chromatin topology or competing for transcription factors.
Adding to these limitations is the restricted experimental framework of most studies. Many studies rely on either single leukemia cell lines or artificial overexpression systems, which do not fully recapitulate the biological complexity of lncRNA–Wnt interactions. In addition, there is a paucity of validation in patient-derived xenografts (PDX) or longitudinal patient cohorts, thus precluding any determination about whether lncRNA-mediated Wnt regulation plays a role in disease initiation, progression, or relapse. Finally, a fundamental unanswered question is whether Wnt-regulating lncRNAs serve as universal modulators across all leukemia subtypes or whether their impact is largely determined by transcriptional and mutational contexts that are subtype-specific.
circRNAs: New players in Wnt pathway regulation
While there is extensive evidence of miRNA and lncRNA involvement in Wnt signaling, circRNA research is still in its infancy. Most evidence of circRNA involvement in Wnt signaling has not been mechanistic in nature and has mainly described the role of circRNAs in regulation. For example, hsa_circ_0121582,106 circ-CBFB,107 and circNPM1108 are reported to regulate Wnt signaling primarily through miRNA sponging, while a wider spectrum of circRNA activity—including protein sequestration, alteration of receptor localization, transcriptional regulation, and translation into peptides—has not been considered in leukemia. Further, while the bulk of circRNA activity is considered to be "predominantly" in the form of miRNA sponging, the complexity more likely arises through additional mechanisms.
Conversely, many studies suggest Wnt activation via increased levels of receptors, such as FZD3 or FZD5, as cited for circ-CBFB107 and circNPM1.108 However, receptor upregulation does not necessarily reflect activation of canonical Wnt/β-catenin signaling. In addition, Frizzled receptors also function in non-canonical Wnt pathways, making it unclear whether the phenotype observed is a direct result of canonical Wnt activation and functionally unrelated to FZD receptor input. Even critical aspects of circRNA biology—i.e., circRNA localization, copy number per cell, and stoichiometric potential to sequester miRNAs—are often not measured, leading to challenges in determining whether proposed Wnt regulatory functions induce physiologically relevant changes.
Furthermore, the field lacks clarity on whether circRNA-mediated Wnt signaling regulation is unique to leukemic stem cells, a feature of drug-resistant subclones, or a general property of malignant hematopoiesis. Determining this is crucial for establishing whether circRNAs can serve as therapeutic targets or biomarkers. Table 2 summarizes the recently recognized effect of ncRNAs on Wnt signaling in leukemia.
Table 2.
ncRNAs affecting Wnt signaling in leukemias
|
ncRNA Type
|
ncRNA name
|
Target(s)/Pathway
|
Effect on Wnt signaling
|
Leukemia type
|
Ref.
|
| miRNA |
miR-155 |
Wnt regulators |
Activates |
ALL, AML |
76,94
|
| miR-370-3p |
PDLIM1 |
Inhibits |
CML |
95
|
| miR-222-3p |
Axin2 |
Activates |
AML |
96
|
| miR-26b-5p |
USP48 |
Inhibits |
AML |
97
|
| miR-1294 |
SOX15 |
Activates |
ALL |
98
|
| miR-301b-3p |
FOXF2 |
Activates |
AML |
109
|
| miR-181a-5p |
Unknown |
Activates |
ALL |
110
|
| miR-221 |
Unknown |
Activates |
ALL |
111
|
| miR-29b |
Sp1/FUT4 |
Activates |
AML |
112
|
| miR-92a |
Unknown |
Activates |
MDS |
113
|
| miR-187-5p |
DKK2 |
Activates |
ALL |
114
|
| miR-136 |
MCL1 |
Activates |
LPS-treated leukemia model |
115
|
| miR-34b |
HSF1 |
Inhibits |
AML |
116
|
| miR-625 |
Unknown |
Activates |
AML |
101
|
| miR-607 |
FZD3 |
Activates |
CLL |
107
|
| LncRNA |
SNHG1 |
miR-489-3p/SOX12 |
Activates |
AML |
100
|
| LINC00909 |
miR-625 |
Activates |
Pediatric AML |
101
|
| DUXAP8 |
Wnt/Glycolysis genes |
Inhibits |
AML |
102
|
| CRNDE |
P-gp/Wnt |
Activates |
AML |
117
|
| HOTAIRM1 |
PFKP |
Activates |
AML |
75
|
| LOC101928834 |
Unknown |
Activates |
MDS |
95
|
| HOTAIR |
Unknown |
Activates |
Mouse leukemia model |
118
|
| LINP1 |
HNF4α/AMPK/WNT5A |
Activates |
AML |
111
|
| circRNA |
hsa_circ_0121582 |
Unknown |
Inhibits |
Leukemia |
106
|
| circ-CBFB |
miR-607/FZD3 |
Activates |
CLL |
107
|
ncRNAs and Wnt-mediated drug resistance in leukemia
While ncRNAs have been implicated in Wnt-driven chemoresistance, there remains a lack of a robust framework to better understand this relationship. The relationship between miR-155 and drug resistance in ALL94 and the relationship between HOTAIRM1 and resistance to cytarabine or daunorubicin in AML103 have been linked to Wnt/β-catenin activation. However, few studies indicate that direct inhibition of Wnt signaling can reverse resistance without concomitant assessment of the ncRNA component. This is a substantial gap in our understanding, as we currently cannot distinguish whether ncRNAs act as primary and upstream drivers of chemoresistance or simply amplify the effects of Wnt-dependent/protected biological pathways. Ultimately, chemoresistance in leukemia arises from a convergence of intersecting mechanisms, including, for example, metabolic adaptations, alterations of mitochondrial dynamics, protective autophagy (as suggested by miR-199a/b-5p119), and the increased expression of efflux transporters; however, most models of resistance, which are currently divorced from Wnt signaling, have failed to integrate these additional layers.
The conflicting descriptions of how Wnt regulates metabolism adds further complexity to the story. For example, HOTAIRM1-mediated Wnt activation is said to increase glycolytic activity,103 while others state the opposite, e.g., DUXAP8-mediated Wnt inhibition leads to reduced glycolysis.102 These conflicting reports suggest that the metabolic functions of Wnt are very context-dependent and don’t apply to all subtypes of leukemia or to treatment status. Finally, there are promising results. For example, circNPM1 is correlated with Adriamycin resistance.108 However, there has been limited or no clinical validation, and very few ncRNA-Wnt signatures have been correlated with patient survival, relapse rate, or treatment response on a wide scale. Fig. 2 illustrates the regulatory functions of ncRNAs on the Wnt signaling pathway. Additionally, Fig. 3 illustrates the interplay between Wnt signaling and the ncRNAs and their role in leukemia.
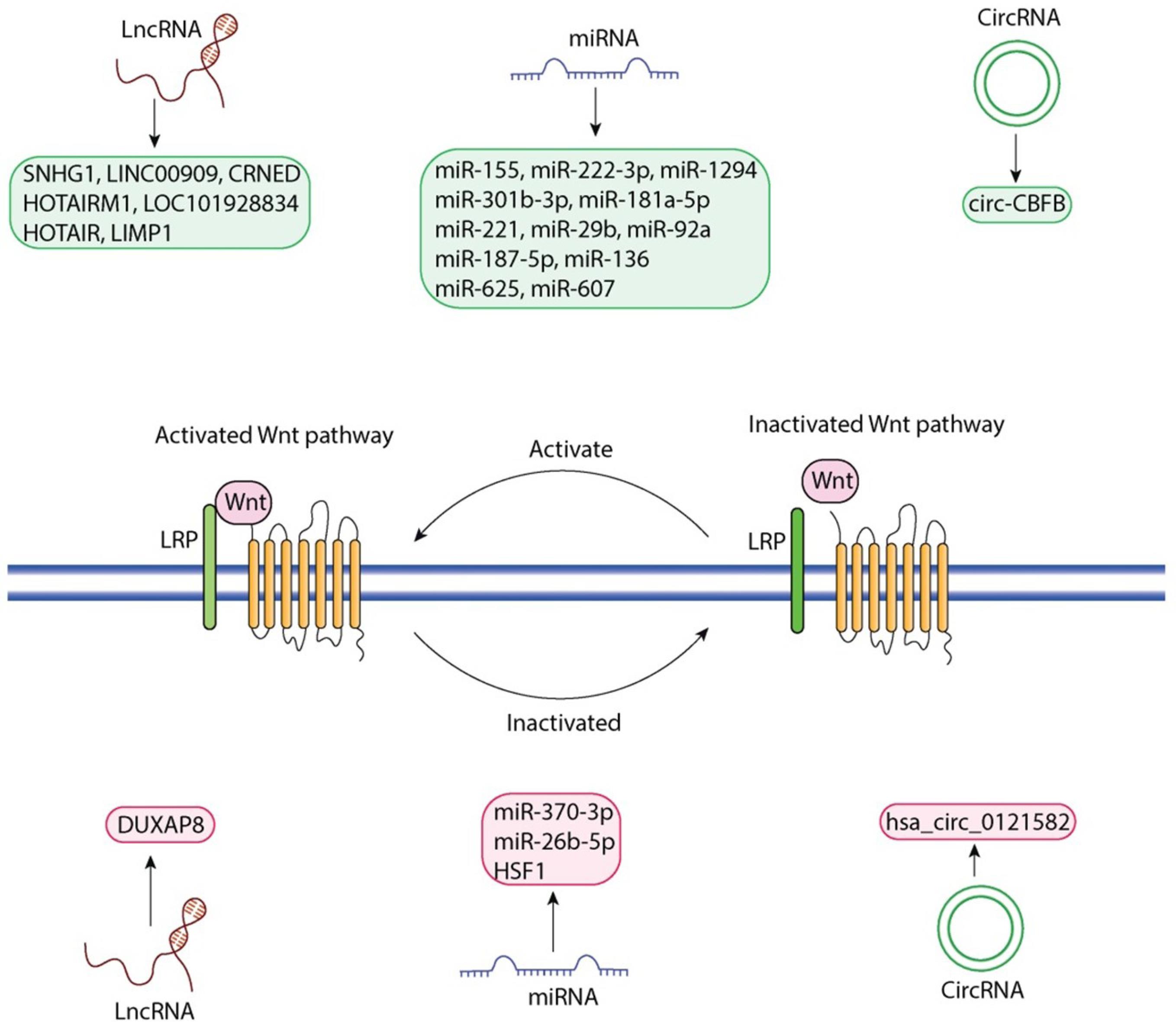
Fig. 2.
Regulatory functions of ncRNAs on the Wnt signaling pathway. This figure demonstrates the multitude of different ncRNA species that influence either the activation or inactivation of the Wnt pathway (left: activated Wnt pathway; right: inactivated Wnt pathway): LncRNAs SNHG1, LINC00909, CRNDE, HOTAIRM1, LOC101928834, HOTAIR, LIMP1, miRNAs miR-155, miR-222-3p, miR-1294, miR-301b-3p, miR-181a-5p, miR-221, miR-29b, miR-92a, miR-187-5p, miR-136, miR-625, miR-607, circRNA circ-CBFB (left: indicated in grey); LncRNA DUXAP8, miRNAs miR-370-3p, miR-26b-5p, HSF1, circRNA hsa_circ_0121582 (right: indicated in grey). In the middle of the figure is the Wnt receptor complex with LRP co-receptors, along with the cyclical activation and inactivation state of Wnt. The figure was generated by Adobe Illustrator 30.0.0.123 software.
.
Regulatory functions of ncRNAs on the Wnt signaling pathway. This figure demonstrates the multitude of different ncRNA species that influence either the activation or inactivation of the Wnt pathway (left: activated Wnt pathway; right: inactivated Wnt pathway): LncRNAs SNHG1, LINC00909, CRNDE, HOTAIRM1, LOC101928834, HOTAIR, LIMP1, miRNAs miR-155, miR-222-3p, miR-1294, miR-301b-3p, miR-181a-5p, miR-221, miR-29b, miR-92a, miR-187-5p, miR-136, miR-625, miR-607, circRNA circ-CBFB (left: indicated in grey); LncRNA DUXAP8, miRNAs miR-370-3p, miR-26b-5p, HSF1, circRNA hsa_circ_0121582 (right: indicated in grey). In the middle of the figure is the Wnt receptor complex with LRP co-receptors, along with the cyclical activation and inactivation state of Wnt. The figure was generated by Adobe Illustrator 30.0.0.123 software.
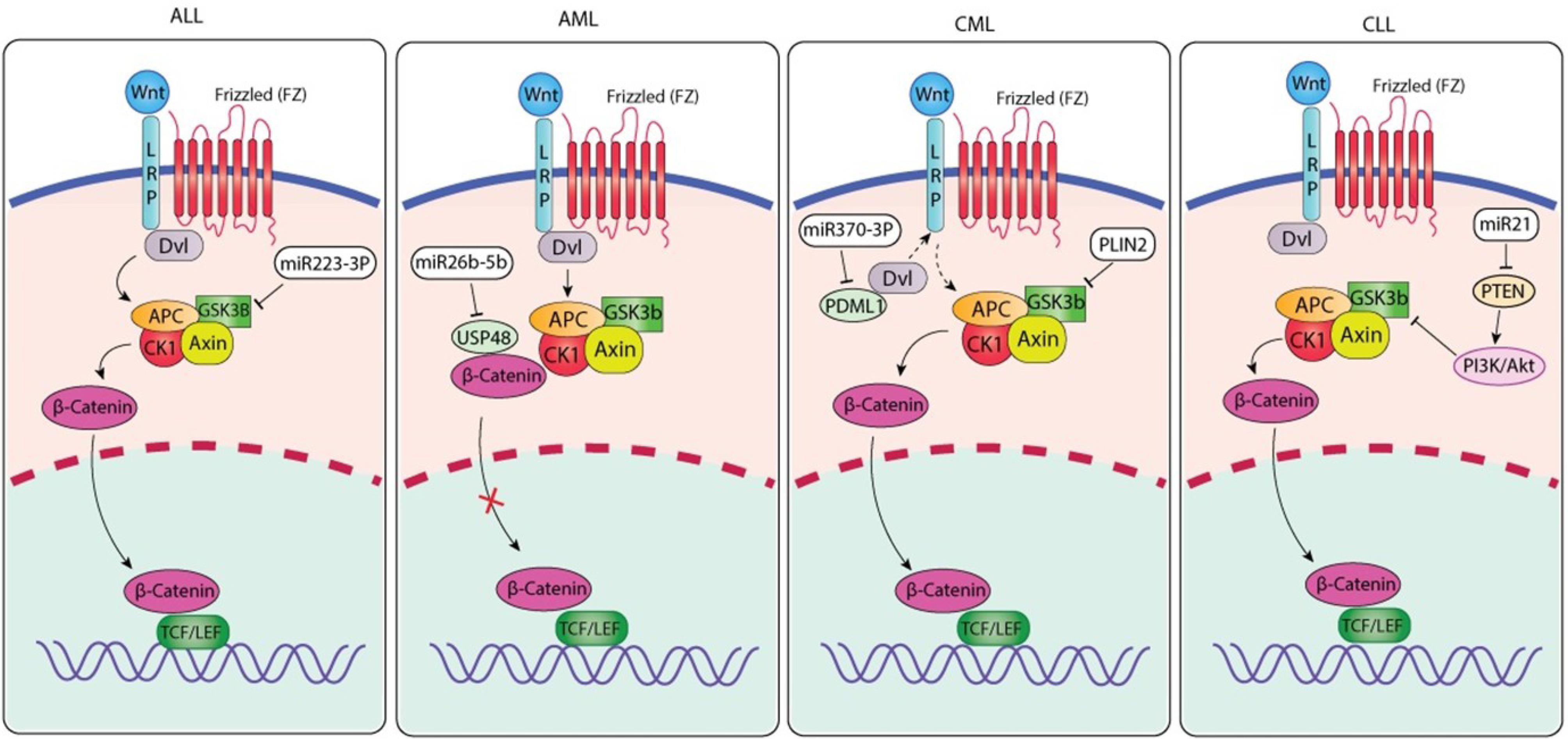
Fig. 3.
ncRNA-Mediated Modulation of Canonical Wnt/β-Catenin Signaling Across Major Leukemia Subtypes. Both Wnt/β-catenin signaling modifiers are modulated by acute lymphoblastic leukemia. Wnt ligands activate and recruit APC/CK1/GSK3β/Axin together with Dvl, which displaces β-catenin from the β-catenin destruction complex (BDC). In this case, miR223-3p represses the expression of GSK3β and reduces the quantity of suitable receptor complexes for β-catenin to form intermediate proteins in the cytoplasm, and upon accumulation in the nucleus, β-catenin co-opts less TCF/LEF transcription factors to activate leukemogenic genes. The acute myeloid leukemia (AML) panel depicts the impairment of the β-catenin destruction complex in AML. Wnt-FZ-LRP signaling impairs the degradation of β-catenin by stimulating the disruption of the APC-CK1-GSK3β-Axin-mediated system. The result of these interactions over time is not only an accumulation of β-catenin but also a subsequent entry into the nucleus. The modulation of Wnt signaling pathway activity is mediated through the repression of USP48 that survives and ultimately inhibits β-catenin stability and activity by identifying regulatory sites. The CML panel indicates that chronic myeloid leukemia has a Wnt signaling (via FZ/LRP) activation of Dvl that reduces the β-catenin destruction complex by Wnt signaling activation. The accumulation and nuclear translocation of β-catenin enable transcription of Wnt-responsive genes. miR-370-3p targets Dvl (intended to indirectly regulate PDML1) by interfering with the upstream signaling strength of Wnt signaling. PLIN2 amplifies β-catenin accumulation to enhance the output of oncogenic Wnt signaling. The CLL panel indicates that chronic lymphocytic leukemia has an alternate means (i.e., ncRNA) of regulating Wnt signaling. Similar to that of CML, Wnt signaling activates Dvl by inhibiting the β-catenin destruction complex, thereby enabling β-catenin accumulation and nuclear translocation. miR-21 inhibits PTEN, and in doing so activates the PI3K/Akt pathway, which subsequently enhances the accumulation and stabilization of β-catenin through the inhibition of GSK3β. The combined action of these signals enhances β-catenin-mediated transcriptional activity and thus promotes the survival and proliferation of leukemic cells. The figure was generated by Adobe Illustrator 30.0.0.123 software.
.
ncRNA-Mediated Modulation of Canonical Wnt/β-Catenin Signaling Across Major Leukemia Subtypes. Both Wnt/β-catenin signaling modifiers are modulated by acute lymphoblastic leukemia. Wnt ligands activate and recruit APC/CK1/GSK3β/Axin together with Dvl, which displaces β-catenin from the β-catenin destruction complex (BDC). In this case, miR223-3p represses the expression of GSK3β and reduces the quantity of suitable receptor complexes for β-catenin to form intermediate proteins in the cytoplasm, and upon accumulation in the nucleus, β-catenin co-opts less TCF/LEF transcription factors to activate leukemogenic genes. The acute myeloid leukemia (AML) panel depicts the impairment of the β-catenin destruction complex in AML. Wnt-FZ-LRP signaling impairs the degradation of β-catenin by stimulating the disruption of the APC-CK1-GSK3β-Axin-mediated system. The result of these interactions over time is not only an accumulation of β-catenin but also a subsequent entry into the nucleus. The modulation of Wnt signaling pathway activity is mediated through the repression of USP48 that survives and ultimately inhibits β-catenin stability and activity by identifying regulatory sites. The CML panel indicates that chronic myeloid leukemia has a Wnt signaling (via FZ/LRP) activation of Dvl that reduces the β-catenin destruction complex by Wnt signaling activation. The accumulation and nuclear translocation of β-catenin enable transcription of Wnt-responsive genes. miR-370-3p targets Dvl (intended to indirectly regulate PDML1) by interfering with the upstream signaling strength of Wnt signaling. PLIN2 amplifies β-catenin accumulation to enhance the output of oncogenic Wnt signaling. The CLL panel indicates that chronic lymphocytic leukemia has an alternate means (i.e., ncRNA) of regulating Wnt signaling. Similar to that of CML, Wnt signaling activates Dvl by inhibiting the β-catenin destruction complex, thereby enabling β-catenin accumulation and nuclear translocation. miR-21 inhibits PTEN, and in doing so activates the PI3K/Akt pathway, which subsequently enhances the accumulation and stabilization of β-catenin through the inhibition of GSK3β. The combined action of these signals enhances β-catenin-mediated transcriptional activity and thus promotes the survival and proliferation of leukemic cells. The figure was generated by Adobe Illustrator 30.0.0.123 software.
Wnt pathway inhibitors
The Wnt/β-catenin signaling pathway is often aberrantly activated in hematologic malignancies, especially in AML and T-ALL. Wnt signaling maintains LSC self-renewal, survival, and chemoresistance, all of which suggest that therapeutic targeting may be possible. Several Wnt pathway inhibitors have emerged from pharmacological studies, presenting a new opportunity for therapeutic intervention in leukemia.
Kamga et al. 120 showed that small-molecule inhibitors of the Wnt/β-catenin signaling axis sensitize AML cells to chemotherapy by disrupting the stabilized microenvironment that supports leukemia. Importantly, restoration of sensitivity was observed when minimal residual disease occurred. Elyamany et al121 reported that cancer cells induce hyperactivation of Wnt signaling and noted that hyperactivation has been linked to deregulation of endogenous Wnt inhibitors, and this effect is more pronounced in older AML patients. Therefore, restoring endogenous Wnt inhibitor function may be beneficial in improving treatment response and outcomes in a high-risk population.
Multiple new compounds show compelling preclinical data supporting Wnt pathway inhibition in leukemia. For example, Wakabayashi et al122 discovered a peptide-type Wnt inhibitor that exhibited potent inhibition of AML cell proliferation. Hadate et al123 discovered “Compound 41” that inhibited Wnt/β-catenin signaling, which inhibited Wnt/β-catenin signaling and induced apoptosis in AML cells. This represents a promising example of a dipeptide Wnt inhibitor with potential as a targeted therapy.
Another challenge in developing Wnt inhibitors has been improving their bioavailability and pharmacokinetic properties. Ai et al124 developed molecules designed to sterically disrupt structural planarity, thereby improving antileukemic activity, and addressing the poor solubility of Wnt inhibitors. This will also be important for advancing effective Wnt inhibitors into clinically viable drugs.
Combination treatment approaches using Wnt inhibition strategies have shown very promising results. Xi and colleagues125 showed that salinomycin, in combination with all-trans retinoic acid (ATRA) was able to activate apoptosis and conversion to a differentiated morphology of AML cells through a decrease in Wnt signaling. Such combinations could have advantages by not only enhancing the apoptotic and differentiating effects of each agent but also targeting multiple leukemogenic mechanisms of action at the same time. Following that work, Qian et al126 also described the impact of combining a BRD4 inhibitor (ACC010) with homoharringtonine. In their study, they demonstrated the synergistic effect their BRD4 inhibitor in combination with homoharringtonine, which suggests that simultaneously targeting the epigenetic regulator and Wnt signaling pathway regulators could be useful, especially in FLT3-ITD-mutant AML.
In T-ALL, Al-Hamaly et al127,128 developed a zebrafish drug screen and identified erlotinib (a well-studied EGFR inhibitor) as a Wnt/β-catenin signaling inhibitor and a suppressor of self-renewal of leukemia cells, which is exciting. Repurposing drugs in this way and identifying Wnt signaling repressors allow clinicians to monitor the clinical effects.
In addition, inhibitors of the upstream regulators of Wnt signaling have been identified with potential therapeutic relevance in preclinical models. In a study published by Mao et al,129 the authors demonstrated that phosphodiesterase 4 inhibition suppressed Wnt signaling and ultimately diminished mitochondrial function and survival of leukemic cells. This also offered a unique metabolic approach to Wnt-targeted therapeutics.
One of the most interesting Wnt inhibitors, BC-2059, was investigated by Soldi et al.130 BC-2059 is able to disrupt the β-catenin/TBL1 complex and inhibit Wnt-dependent gene transcription. Given its specific mechanism of action, BC-2059 can target downstream transcriptional outputs while circumventing primary upstream signaling components and thus likely reducing potential off-target effects. Table 3 summarizes the Wnt pathway inhibitors used in leukemias.
Table 3.
Wnt/β-catenin signaling pathway inhibitors used in leukemias
|
Compound
|
Target
|
Leukemia type
|
Mechanism
|
Notes
|
Ref.
|
| Small Molecule Inhibitors |
Wnt/β-catenin |
AML |
Enhances chemosensitivity |
Disrupts microenvironmental support |
120
|
| -- |
Endogenous inhibitors |
AML (elderly) |
Restores regulation |
Targets pathway deregulation |
121
|
| Dipeptide Inhibitor |
Wnt/β-catenin |
AML |
Suppresses proliferation |
Novel small molecule |
122
|
| Compound #41 |
Wnt/β-catenin |
AML |
Inhibits signaling, induces apoptosis |
High efficacy in vitro |
123
|
| Improved Inhibitor |
Wnt/β-catenin |
AML |
Increased solubility and potency |
Structure modification |
124
|
| Salinomycin + ATRA |
Wnt/β-catenin |
AML |
Induces apoptosis/differentiation |
Combination therapy |
125
|
| Erlotinib |
Wnt/β-catenin |
T-ALL |
Inhibits self-renewal |
Identified via zebrafish screening |
127,128
|
| PDE4 Inhibitor |
Wnt/β-catenin (via mitochondria) |
AML |
Impaired energy metabolism |
Targeting metabolic axis |
129
|
| ACC010 + HHT |
BRD4/Wnt pathway |
AML (FLT3-ITD) |
Synergistic inhibition |
Epigenetic + Wnt targeting |
126
|
| BC-2059 |
β-catenin/TBL1 complex |
Multiple cancers |
Blocks transcription |
Highly specific inhibitor |
130
|
Therapeutic potentials
A recent expansion in the awareness and role of ncRNA in the treatment of disease has substantially changed how ncRNA is viewed and used in disease progression and as a possible therapeutic target. With the discovery of new roles for miRNA, lncRNA, circRNA, etc., as well as the existence of preclinical evidence demonstrating the use of miRNA mimics and antisense oligonucleotide technologies that are capable of modulating and affecting the genetic networks associated with the pathogenesis of cancer and other diseases, the amount of data supporting the potential of these drug delivery methods is growing rapidly.131-133 Nonetheless, transferring the preclinical success of these ncRNA technologies into clinical practice will require overcoming major delivery, stability, distribution, and immunogenicity obstacles. Currently available strategies to conquer these obstacles include the use of engineered chemically stabilized miRNA mimic or antisense oligonucleotides to complement physiologic miRNA function and/or prevent the activity of oncogenic miRNA. Many synthetic miRNA mimic have demonstrated effectiveness against a wide variety of solid tumors, hematologic malignancies, and inflammatory diseases through the restoration of physiologically functioning gene regulatory circuits.131,132 However, despite compelling biological rationales, administration of miRNA mimic into the circulation of an individual is complicated by biological processes such as nucleic acid degeneration in plasma and the rapid clearance of miRNA mimics from circulation via the renal route. As a result, a great deal of research has been done to develop effective delivery systems for ncRNA-based products.134,135
The use of engineered carriers to protect and to promote delivery of ncRNA therapeutics into the intended site of action is increasingly being developed. For example, while lipid nanoparticles (LNPs) have provided a stable platform for approved RNA-based drugs, LNPs are also being modified for enhanced stability in serum and improved cellular uptake and endosomal escape efficiency.136,137 Recent advances in engineering, including ionizable lipids, pH-responsive polymers, and fusogenic peptides, have been designed to address entrapment during transport within an endosome. These advances have been instrumental in making RNA expression in cytosols safer.137 In addition to engineered LNPs, several teams are currently working toward developing an exosome-based method of delivering RNA, which would have several advantages over existing LNP-based RNA delivery. The use of engineered exosomes to transport miRNA mimics or inhibitors has demonstrated excellent potential to selectively inhibit tumor growth or induce apoptosis in tumor cells. However, scalability of manufacturing these exosome-based delivery systems remains a barrier to successful clinical translation.136,138
An alternative to traditional methods of delivering various products via lipid systems is the use of nanoparticles made with polymers and polysaccharides. One such example is chitosan. This is naturally occurring polymer that has excellent biocompatibility and stability, and can be engineered to specifically target cancerous/inflamed tissues with ncRNA (such as miRNAs and siRNAs).139 Other approaches with RNA nanostructures exhibiting inherent stability and modularity have shown potential in several diseases, such as hepatocellular carcinoma (HCC), where programmable RNA scaffolds with multiple therapeutic modules can affect oncogenic signaling pathways at once 134. Likewise, for treatment of cardiovascular and metabolic disease models, peptides, antibodies, or ligand-receptor systems are being used to optimize targeted delivery methods that result in specific uptake of ncRNA therapies by cells located in vascular tissues, macrophages, or cardiomyocytes.140,141
The use of circRNA-targeted therapies is an area showing rapid progress. circRNA have many functions, including acting as a sponges, scaffolds, and transcription regulators, and the fact that circRNAs have a covalently closed structure makes them incredibly stable in vivo. There are several strategies to influence circRNAs in a therapeutic setting, such as by creating oligonucleotides that bind to pathogenic circRNAs and cause them to degrade, or alternatively developing synthetic circRNAs that function as highly competitive inhibitors or protective RNA sponges. 136,142 While the technologies to create these products are currently in development, the high stability and tissue-specific properties of circRNAs greatly enhance their potential to be future drug targets.136,142
At the same time that therapeutic applications are being developed, there is a significant effort underway to evaluate circulating non-coding RNAs as potential minimally invasive biomarkers for diagnosis, prognosis, and monitoring of treatment outcomes. The high stability of ncRNAs in biofluids (blood, saliva, and CSF) is due to the fact that they are protected from degradation by being contained within extracellular vesicles, lipoproteins, or ribonucleoprotein complexes.142,143 Many studies from different patient cohorts have identified the unique profiles of circulating ncRNA that are associated with a variety of clinical conditions, including cancer progression, treatment resistance, cardiovascular disease severity, and neurological disorders. For example, various miRNA panels in plasma have been found to have diagnostic accuracy in large cancer populations and are superior to conventional protein biomarkers for both sensitivity and the ability to distinguish different stages of disease.131,144 Similarly, miRNAs isolated from exosomes have been validated in cohort studies as prognostic markers in lung cancer and correlated with tumor burden and therapeutic response.144
The potential for miRNA to be developed into clinically relevant biomarkers is demonstrated via numerous clinical trials targeting the assessment of circulating miRNA to determine their usefulness as an indicator of therapeutic efficacy and risk of relapse when evaluating patients diagnosed with hepatocellular carcinoma, breast carcinoma, and colorectal carcinoma using miR-21, miR-122, and miR-200 family members as predictive biomarkers to response to therapy. CircRNAs are at the same level of developmental activity as miRNAs; researchers are beginning to use circRNAs to create biomarker panels to provide insight into the underlying biology of the tumors and the risk of recurrence and relapse to predict survival. CircRNAs have shown promise in the field of neurology, where researchers have studied unique profiles of specific circRNAs in the plasma and cerebrospinal fluid of patients diagnosed with temporal lobe epilepsy. Researchers have documented differences in circRNA levels in the blood of patients with different cancers and cardiovascular diseases. Together with their stability, these characteristics make them highly desirable as biomarkers for clinical use. Therefore, researchers must incorporate ncRNA profiling into clinical trial protocols to further improve the validation of ncRNA biomarkers and the establishment of standardized quantification methods for these markers.
Conclusion and future perspectives
Consequently, based on the cellular environment and the molecules present or co-expressed with the genes, as well as the form of leukemia, these genes (onco/tumor suppressor genes) may may promote or inhibit leukemia development. The modulation of Wnt signaling in lymphocytic leukemia represents a potential target for new therapies. Therefore, they can act as either oncogenes or tumor suppressor genes depending on the cellular context, the specific molecular partners that are expressed in conjunction with them, and the relevant leukemia subtype. Their ability to modulate Wnt signaling is an indicator of their potential utility as potential targets of therapeutic intervention in the treatment of leukemia.
Research priorities will focus on developing high-resolution maps of ncRNA-Wnt signaling interactions through the use of high-throughput transcriptomic, epigenomic, proteomic, and single-cell analyses in order to identify subtype-specific circuits, as well as discover novel regulatory ncRNAs. In addition, it is important to explore how multiple ncRNAs interact to modulate both canonical and non-canonical Wnt signaling (collectively) in order to understand the combined regulatory impact of ncRNAs on the Wnt signaling pathway. Functional studies examining the influence of ncRNA-Wnt interactions on stemness, clonal evolution in response to treatment, and progression of leukemia should include the use of CRISPR perturbations, antisense oligonucleotides, RNAi (RNA interference), and relevant in vivo models (e.g., PDX and organoids). In particular, ncRNAs involved in modulating non-canonical Wnt signaling pathways have not yet been characterized in leukemic systems and may provide insight into additional mechanisms of resistance or disease progression.
When used therapeutically together, Wnt pathway enzymes (such as tankyrase and porcupine) inhibitors and ncRNA-directed methods such as miRNA mimics, antagomirs, lncRNA silencers, and circRNA silencers are approaches to target cancer-causing signal loops, particularly those that maintain LSCs. Also, the availability of new delivery methods for RNA (lipid nanoparticles and exosomes) will aid in making therapeutic decisions more accurately. Lastly, circulating ncRNAs (particularly circRNA) are expected to be used as noninvasive predictive biomarkers for diagnostics, risk stratification, treatment monitoring, and predicting relapse. Therefore, profiling of clinical data for patients will aid in making therapeutic decisions more accurately by continuously profiling the ncRNA signature for each patient and determining the level of Wnt activity, as these will be critical components in advancing personalized therapeutics through better integration of patient-specific ncRNA and Wnt activity data into PDX and organoid platforms.
Review Highlights
What is the current knowledge?
-
The Wnt signaling pathway functions as a major regulator of hematopoietic stem cell self-renewal, and significantly deregulated in several leukemia types (e.g., AML, CML, ALL, and CLL) in the context of leukemogenesis, disease progression, and resistance to drug therapies.
-
ncRNAs, such as miRNAs, lncRNAs, and circRNAs, are important regulators of gene expression and are increasingly understood to be important players in hematological malignancies.
-
Current evidence has suggested ncRNAs can directly or indirectly modulate Wnt pathway components impacting leukemic cell proliferation, survival, and response to therapy.
What is new here?
-
In summary, this review provides the most extensive review to date of the various ncRNAs (miRNAs, lncRNAs, circRNAs) that regulate (and are regulated by) Wnt signaling in leukemia.
-
The authors highlight the bidirectional nature of the ncRNA-Wnt signaling interactions since ncRNAs regulate Wnt signaling, while activation of the Wnt pathway can also affect these ncRNAs, and likely creates feedback loops that maintain leukemic phenotypes.
-
This manuscript categorizes and describes how ncRNAs regulate the canonical Wnt and non-canonical Wnt signaling components, regulation of leukemic stem cell biology, and aid in drug resistance in leukemia.
-
The authors highlight ncRNA-Wnt interactions as potential biomarkers for diagnosis and prognosis, and therapeutic targets suggesting prospective RNA-based and combination treatment approaches for leukemia.
Competing Interests
The authors have no relevant financial or non-financial interests to disclose.
Declaration of AI-assisted Tools in the Writing Procedure
The authors used ChatGPT version 5 for language edition of the paper. The authors reviewed the text for accuracy and take full responsibility for the final content.
Data Availability Statement
Data are available on request from the authors.
Ethical Approval
Not applicable.
Acknowledgements
This work was done as a part of M.Sc thesis for Reza Biglari. Authors would acknowledge department of biochemistry and Clinical laboratories at Tabriz University of Medical Sciences, Tabriz, Iran.
References
-
Wachter F, Pikman Y. Pathophysiology of acute myeloid leukemia. Acta Haematol 2024. 147: 229-46. doi: 10.1159/000536152.
- Bouligny IM, Maher KR, Grant S. Mechanisms of myeloid leukemogenesis: current perspectives and therapeutic objectives. Blood Rev 2023; 57:100996. doi: 10.1016/j.blre.2022.100996 [Crossref] [ Google Scholar]
- Ley TJ, Miller C, Ding L, Raphael BJ, Mungall AJ, Robertson A. Genomic and epigenomic landscapes of adult de novo acute myeloid leukemia. N Engl J Med 2013; 368:2059-74. doi: 10.1056/NEJMoa1301689 [Crossref] [ Google Scholar]
- Armstrong SA, Look AT. Molecular genetics of acute lymphoblastic leukemia. J Clin Oncol 2005; 23:6306-15. doi: 10.1200/jco.2005.05.047 [Crossref] [ Google Scholar]
- Rinaldi I, Winston K. Chronic myeloid leukemia, from pathophysiology to treatment-free remission: a narrative literature review. J Blood Med 2023; 14:261-77. doi: 10.2147/jbm.S382090 [Crossref] [ Google Scholar]
- Arber DA, Orazi A, Hasserjian R, Thiele J, Borowitz MJ, Le Beau MM. The 2016 revision to the World Health Organization classification of myeloid neoplasms and acute leukemia. Blood 2016; 127:2391-405. doi: 10.1182/blood-2016-03-643544 [Crossref] [ Google Scholar]
- Ruan Y, Kim HN, Ogana H, Kim YM. Wnt signaling in leukemia and its bone marrow microenvironment. Int J Mol Sci 2020; 21:6247. doi: 10.3390/ijms21176247 [Crossref] [ Google Scholar]
- Soares-Lima SC, Pombo-de-Oliveira MS, Carneiro FR. The multiple ways Wnt signaling contributes to acute leukemia pathogenesis. J Leukoc Biol 2020; 108:1081-99. doi: 10.1002/jlb.2mr0420-707r [Crossref] [ Google Scholar]
- Láinez-González D, Alonso-Aguado AB, Alonso-Dominguez JM. Understanding the Wnt signaling pathway in acute myeloid leukemia stem cells: a feasible key against relapses. Biology (Basel) 2023; 12:683. doi: 10.3390/biology12050683 [Crossref] [ Google Scholar]
- Bhattacharya M, Sharma AR, Sharma G, Patra BC, Lee SS, Chakraborty C. Interaction between miRNAs and signaling cascades of Wnt pathway in chronic lymphocytic leukemia. J Cell Biochem 2020; 121:4654-66. doi: 10.1002/jcb.29683 [Crossref] [ Google Scholar]
- Pehlivan M, Soyoz M, Cerci B, Coven HIK, Yuce Z, Sercan HO. [sFRP1 expression induces miRNAs that modulate Wnt signaling in chronic myeloid leukemia cells]. Mol Biol (Mosk) 2020; 54:626-33. doi: 10.31857/s002689842004014x [Crossref] [ Google Scholar]
- Sheng XF, Hong LL, Li H, Huang FY, Wen Q, Zhuang HF. Long non-coding RNA MALAT1 modulate cell migration, proliferation and apoptosis by sponging microRNA-146a to regulate CXCR4 expression in acute myeloid leukemia. Hematology 2021; 26:43-52. doi: 10.1080/16078454.2020.1867781 [Crossref] [ Google Scholar]
- Chen JJ, Lei P, Zhou M. hsa_circ_0121582 inhibits leukemia growth by dampening Wnt/β-catenin signaling. Clin Transl Oncol 2020; 22:2293-302. doi: 10.1007/s12094-020-02377-9 [Crossref] [ Google Scholar]
- Shi Z, Li J, Ding J, Zhang Y, Min W, Zhu Y. ADAR1 is required for acute myeloid leukemia cell survival by modulating post-transcriptional Wnt signaling through impairing miRNA biogenesis. Leukemia 2025; 39:599-613. doi: 10.1038/s41375-024-02500-7 [Crossref] [ Google Scholar]
- Yu S, Han R, Gan R. The Wnt/β-catenin signalling pathway in haematological neoplasms. Biomark Res 2022; 10:74. doi: 10.1186/s40364-022-00418-9 [Crossref] [ Google Scholar]
- Krimpenfort RA, Nethe M. Canonical Wnt: a safeguard and threat for erythropoiesis. Blood Adv 2021; 5:3726-35. doi: 10.1182/bloodadvances.2021004845 [Crossref] [ Google Scholar]
- Evangelisti C, Chiarini F, Cappellini A, Paganelli F, Fini M, Santi S. Targeting Wnt/β-catenin and PI3K/Akt/mTOR pathways in T-cell acute lymphoblastic leukemia. J Cell Physiol 2020; 235:5413-28. doi: 10.1002/jcp.29429 [Crossref] [ Google Scholar]
- Wagstaff M, Sevim O, Goff A, Raynor M, Park H, Mancini EJ. β-Catenin interacts with canonical RBPs including MSI2 to associate with a Wnt signalling mRNA network in myeloid leukaemia cells. Oncogene 2025; 44:2490-503. doi: 10.1038/s41388-025-03415-y [Crossref] [ Google Scholar]
- Marr AR, Halpin M, Corbin DL, Asemelash Y, Sher S, Gordon BK. The multi-CDK inhibitor dinaciclib reverses bromo- and extra-terminal domain (BET) inhibitor resistance in acute myeloid leukemia via inhibition of Wnt/β-catenin signaling. Exp Hematol Oncol 2024; 13:27. doi: 10.1186/s40164-024-00483-w [Crossref] [ Google Scholar]
- Nakano K, Chihara Y, Kobayashi S, Iwanaga M, Utsunomiya A, Watanabe T. Overexpression of aberrant Wnt5a and its effect on acquisition of malignant phenotypes in adult T-cell leukemia/lymphoma (ATL) cells. Sci Rep 2021; 11:4114. doi: 10.1038/s41598-021-83613-2 [Crossref] [ Google Scholar]
- Caliskan C, Yuce Z, Ogun Sercan H. Dvl proteins regulate SMAD1, AHR, mTOR, BRD7 protein expression while differentially regulating canonical and non-canonical Wnt signaling pathways in CML cell lines. Gene 2023; 854:147109. doi: 10.1016/j.gene.2022.147109 [Crossref] [ Google Scholar]
- Bhat AA, Younes SN, Raza SS, Zarif L, Nisar S, Ahmed I. Role of non-coding RNA networks in leukemia progression, metastasis and drug resistance. Mol Cancer 2020; 19:57. doi: 10.1186/s12943-020-01175-9 [Crossref] [ Google Scholar]
- Liu Y, Sun P, Zhao Y, Liu B. The role of long non-coding RNAs and downstream signaling pathways in leukemia progression. Hematol Oncol 2021; 39:27-40. doi: 10.1002/hon.2776 [Crossref] [ Google Scholar]
- Anelli L, Zagaria A, Specchia G, Musto P, Albano F. Dysregulation of miRNA in leukemia: exploiting miRNA expression profiles as biomarkers. Int J Mol Sci 2021; 22:7156. doi: 10.3390/ijms22137156 [Crossref] [ Google Scholar]
- Javandoost E, Firoozi-Majd E, Rostamian H, Khakpoor-Koosheh M, Mirzaei HR. Role of microRNAs in chronic lymphocytic leukemia pathogenesis. Curr Med Chem 2020; 27:282-97. doi: 10.2174/0929867326666190911114842 [Crossref] [ Google Scholar]
- Zhang R, Khare P, Banerjee P, Ivan C, Schneider S, Barbaglio F. The DLEU2/miR-15a/miR-16-1 cluster shapes the immune microenvironment of chronic lymphocytic leukemia. Blood Cancer J 2024; 14:168. doi: 10.1038/s41408-024-01142-3 [Crossref] [ Google Scholar]
- Guo B, Qin R, Chen JJ, Pan W, Lu XC. MicroRNA-125b accelerates and promotes PML-RARa-driven murine acute promyelocytic leukemia. Biomed Environ Sci 2022; 35:485-93. doi: 10.3967/bes2022.067 [Crossref] [ Google Scholar]
- Philippe C, Jaud M, Féral K, Gay A, Van Den Berghe L, Farce M. Pivotal role of the endoplasmic reticulum stress-related XBP1s/miR-22/SIRT1 axis in acute myeloid leukemia apoptosis and response to chemotherapy. Leukemia 2024; 38:1764-76. doi: 10.1038/s41375-024-02321-8 [Crossref] [ Google Scholar]
- Xia J, Bu C, Zhang B, Wang X, Chen Y, Li T. The emerging role of microRNA-22 in the leukemia: experimental and clinical implications. Mol Biol Rep 2023; 51:12. doi: 10.1007/s11033-023-08922-3 [Crossref] [ Google Scholar]
- Pang B, Mao H, Wang J, Yang W. MiR-185-5p suppresses acute myeloid leukemia by inhibiting GPX1. Microvasc Res 2022; 140:104296. doi: 10.1016/j.mvr.2021.104296 [Crossref] [ Google Scholar]
- Wen J, Chen Y, Liao C, Ma X, Wang M, Li Q. Engineered mesenchymal stem cell exosomes loaded with miR-34c-5p selectively promote eradication of acute myeloid leukemia stem cells. Cancer Lett 2023; 575:216407. doi: 10.1016/j.canlet.2023.216407 [Crossref] [ Google Scholar]
- Caserta C, Nucera S, Barcella M, Fazio G, Naldini MM, Pagani R. miR-126 identifies a quiescent and chemo-resistant human B-ALL cell subset that correlates with minimal residual disease. Leukemia 2023; 37:1994-2005. doi: 10.1038/s41375-023-02009-5 [Crossref] [ Google Scholar]
- Qiao J, Zhao D, Nguyen LX, Chen F, Liang C, Estrella K. Targeting miR-126 in Ph + acute lymphoblastic leukemia. Leukemia 2023; 37:1540-4. doi: 10.1038/s41375-023-01933-w [Crossref] [ Google Scholar]
- Farzaneh M, Najafi S, Sheykhi-Sabzehpoush M, Nezhad Dehbashi F, Anbiyaee O, Nasrolahi A. The stem cell-specific long non-coding RNAs in leukemia. Clin Transl Oncol 2023; 25:345-51. doi: 10.1007/s12094-022-02952-2 [Crossref] [ Google Scholar]
- Izadirad M, Jafari L, James AR, Unfried JP, Wu ZX, Chen ZS. Long noncoding RNAs have pivotal roles in chemoresistance of acute myeloid leukemia. Drug Discov Today 2021; 26:1735-43. doi: 10.1016/j.drudis.2021.03.017 [Crossref] [ Google Scholar]
- Dieter C, Lourenco ED, Lemos NE. Association of long non-coding RNA and leukemia: a systematic review. Gene 2020; 735:144405. doi: 10.1016/j.gene.2020.144405 [Crossref] [ Google Scholar]
- Sabaghi F, Yousefi Sadat S, Mirsaeedi Z, Salahi A, Vazifehshenas S, Zahmat Kesh N. The role of long noncoding RNAs in progression of leukemia: based on chromosomal location. Microrna 2024; 13:14-32. doi: 10.2174/0122115366265540231201065341 [Crossref] [ Google Scholar]
- Shi H, Gao L, Zhang W, Jiang M. Long non-coding RNAs regulate treatment outcome in leukemia: what have we learnt recently?. Cancer Med 2023; 12:13966-77. doi: 10.1002/cam4.6027 [Crossref] [ Google Scholar]
- Rudich A, Garzon R, Dorrance A. Non-coding RNAs are implicit in chronic myeloid leukemia therapy resistance. Int J Mol Sci 2022; 23:12271. doi: 10.3390/ijms232012271 [Crossref] [ Google Scholar]
- Liu Y, Zhu XY, Liao LL, Zhang ZH, Huang TS, Zhang L. Silencing LINC00987 ameliorates Adriamycin resistance of acute myeloid leukemia via miR-4458/HMGA2 axis. Biol Direct 2024; 19:49. doi: 10.1186/s13062-024-00490-1 [Crossref] [ Google Scholar]
- Jiang Z, Liu T, Wang Y, Li J, Guo L. Effect of lncRNA XIST on acute myeloid leukemia cells via miR-142-5p-PFKP axis. Hematology 2024; 29:2306444. doi: 10.1080/16078454.2024.2306444 [Crossref] [ Google Scholar]
- Zhao Y, Wang W, Guan C, Hu Z, Liu L, Li W. Long noncoding RNA HOTAIRM1 in human cancers. Clin Chim Acta 2020; 511:255-9. doi: 10.1016/j.cca.2020.10.011 [Crossref] [ Google Scholar]
- Ghahramani Almanghadim H, Karimi B, Poursalehi N, Sanavandi M, Atefi Pourfardin S, Ghaedi K. The biological role of lncRNAs in the acute lymphocytic leukemia: an updated review. Gene 2024; 898:148074. doi: 10.1016/j.gene.2023.148074 [Crossref] [ Google Scholar]
- Jalilivand S, Izadirad M, Vazifeh Shiran N, Gharehbaghian A, Naserian S. The effect of bone marrow mesenchymal stromal cell exosomes on acute myeloid leukemia's biological functions: a focus on the potential role of LncRNAs. Clin Exp Med 2024; 24:108. doi: 10.1007/s10238-024-01364-6 [Crossref] [ Google Scholar]
- Li Q, Ren X, Wang Y, Xin X. CircRNA: a rising star in leukemia. PeerJ 2023; 11:e15577. doi: 10.7717/peerj.15577 [Crossref] [ Google Scholar]
- Singh V, Uddin MH, Zonder JA, Azmi AS, Balasubramanian SK. Circular RNAs in acute myeloid leukemia. Mol Cancer 2021; 20:149. doi: 10.1186/s12943-021-01446-z [Crossref] [ Google Scholar]
- Deng W, Chao R, Zhu S. Emerging roles of circRNAs in leukemia and the clinical prospects: an update. Immun Inflamm Dis 2023; 11:e725. doi: 10.1002/iid3.725 [Crossref] [ Google Scholar]
- Zhou Q, Shu X, Chai Y, Liu W, Li Z, Xi Y. The non-coding competing endogenous RNAs in acute myeloid leukemia: biological and clinical implications. Biomed Pharmacother 2023; 163:114807. doi: 10.1016/j.biopha.2023.114807 [Crossref] [ Google Scholar]
- Yeh SC, Cheong FJF, Tay Y. Circular RNAs and untranslated regions in acute myeloid leukemia. Int J Mol Sci 2023; 24:3215. doi: 10.3390/ijms24043215 [Crossref] [ Google Scholar]
- Gutierrez-Camino A, Caron M, Richer C, Fuchs C, Illarregi U, Poncelet L. CircRNAome of childhood acute lymphoblastic leukemia: deciphering subtype-specific expression profiles and involvement in TCF3::PBX1 ALL. Int J Mol Sci 2024; 25:1477. doi: 10.3390/ijms25031477 [Crossref] [ Google Scholar]
- das Chagas PF, de Sousa GR, Kodama MH, de Biagi Junior CA, Yunes JA, Brandalise SR. Ultraconserved long non-coding RNA uc112 is highly expressed in childhood T versus B-cell acute lymphoblastic leukemia. Hematol Transfus Cell Ther 2021; 43:28-34. doi: 10.1016/j.htct.2019.12.003 [Crossref] [ Google Scholar]
- Shi J, Ding W, Lu H. Identification of long non-coding RNA SNHG family as promising prognostic biomarkers in acute myeloid leukemia. Onco Targets Ther 2020; 13:8441-50. doi: 10.2147/ott.S265853 [Crossref] [ Google Scholar]
- Yan H, Zhang DY, Li X, Yuan XQ, Yang YL, Zhu KW. Long non-coding RNA GAS5 polymorphism predicts a poor prognosis of acute myeloid leukemia in Chinese patients via affecting hematopoietic reconstitution. Leuk Lymphoma 2017; 58:1948-57. doi: 10.1080/10428194.2016.1266626 [Crossref] [ Google Scholar]
- Huang JL, Liu W, Tian LH, Chai TT, Liu Y, Zhang F. Upregulation of long non-coding RNA MALAT-1 confers poor prognosis and influences cell proliferation and apoptosis in acute monocytic leukemia. Oncol Rep 2017; 38:1353-62. doi: 10.3892/or.2017.5802 [Crossref] [ Google Scholar]
- Pouyanrad S, Rahgozar S, Ghodousi ES. Dysregulation of miR-335-3p, targeted by NEAT1 and MALAT1 long non-coding RNAs, is associated with poor prognosis in childhood acute lymphoblastic leukemia. Gene 2019; 692:35-43. doi: 10.1016/j.gene.2019.01.003 [Crossref] [ Google Scholar]
- Guan J, Liu P, Wang A, Wang B. Long non-coding RNA ZEB2-AS1 affects cell proliferation and apoptosis via the miR-122-5p/PLK1 axis in acute myeloid leukemia. Int J Mol Med 2020; 46:1490-500. doi: 10.3892/ijmm.2020.4683 [Crossref] [ Google Scholar]
- Luo W, Yu H, Zou X, Ni X, Wei J. Long non-coding RNA taurine-upregulated gene 1 correlates with unfavorable prognosis in patients with refractory or relapsed acute myeloid leukemia treated by purine analogue based chemotherapy regimens. Cancer Biomark 2018; 23:485-94. doi: 10.3233/cbm-181405 [Crossref] [ Google Scholar]
- Wang X, Zhang L, Zhao F, Xu R, Jiang J, Zhang C. Long non-coding RNA taurine-upregulated gene 1 correlates with poor prognosis, induces cell proliferation, and represses cell apoptosis via targeting aurora kinase A in adult acute myeloid leukemia. Ann Hematol 2018; 97:1375-89. doi: 10.1007/s00277-018-3315-8 [Crossref] [ Google Scholar]
- Qin J, Bao H, Li H. Correlation of long non-coding RNA taurine-upregulated gene 1 with disease conditions and prognosis, as well as its effect on cell activities in acute myeloid leukemia. Cancer Biomark 2018; 23:569-77. doi: 10.3233/cbm-181834 [Crossref] [ Google Scholar]
- Huang R, Liao X, Wang X, Li Q. Comprehensive investigation of the clinical significance of long non-coding RNA HOXA-AS2 in acute myeloid leukemia using genome-wide RNA sequencing dataset. J Cancer 2021; 12:2151-64. doi: 10.7150/jca.48045 [Crossref] [ Google Scholar]
- Huang HH, Chen FY, Chou WC, Hou HA, Ko BS, Lin CT. Long non-coding RNA HOXB-AS3 promotes myeloid cell proliferation and its higher expression is an adverse prognostic marker in patients with acute myeloid leukemia and myelodysplastic syndrome. BMC Cancer 2019; 19:617. doi: 10.1186/s12885-019-5822-y [Crossref] [ Google Scholar]
- Papaioannou D, Petri A, Dovey OM, Terreri S, Wang E, Collins FA. Publisher correction: the long non-coding RNA HOXB-AS3 regulates ribosomal RNA transcription in NPM1-mutated acute myeloid leukemia. Nat Commun 2020; 11:204. doi: 10.1038/s41467-019-13969-7 [Crossref] [ Google Scholar]
- Papaioannou D, Petri A, Dovey OM, Terreri S, Wang E, Collins FA. Author correction: the long non-coding RNA HOXB-AS3 regulates ribosomal RNA transcription in NPM1-mutated acute myeloid leukemia. Nat Commun 2020; 11:3878. doi: 10.1038/s41467-020-17746-9 [Crossref] [ Google Scholar]
- Cui C, Wang Y, Gong W, He H, Zhang H, Shi W. Long non-coding RNA LINC00152 regulates self-renewal of leukemia stem cells and induces chemo-resistance in acute myeloid leukemia. Front Oncol 2021; 11:694021. doi: 10.3389/fonc.2021.694021 [Crossref] [ Google Scholar]
- Wang J, Liu ZH, Yu LJ. Long non-coding RNA LINC00641 promotes cell growth and migration through modulating miR-378a/ZBTB20 axis in acute myeloid leukemia. Eur Rev Med Pharmacol Sci 2019; 23:7498-509. doi: 10.26355/eurrev_201909_18864 [Crossref] [ Google Scholar]
- Liu Y, Gao X, Tian X. High expression of long intergenic non-coding RNA LINC00662 contributes to malignant growth of acute myeloid leukemia cells by upregulating ROCK1 via sponging microRNA-340-5p. Eur J Pharmacol 2019; 859:172535. doi: 10.1016/j.ejphar.2019.172535 [Crossref] [ Google Scholar]
- Chen B, Li Y, Nie Y, Tang A, Zhou Q. Long non-coding RNA LINC01268 promotes cell growth and inhibits cell apoptosis by modulating miR-217/SOS1 axis in acute myeloid leukemia. Braz J Med Biol Res 2020; 53:e9299. doi: 10.1590/1414-431x20209299 [Crossref] [ Google Scholar]
- Chen W, Gan DY. [Effect of long non-coding RNA LINC01268 on the malignant biological behaviors of acute myeloid leukemia cells]. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2023; 31:1608-16. doi: 10.19746/j.cnki.issn.1009-2137.2023.06.002 [Crossref] [ Google Scholar]
- Wang Y, Li Y, Song HQ, Sun GW. Long non-coding RNA LINC00899 as a novel serum biomarker for diagnosis and prognosis prediction of acute myeloid leukemia. Eur Rev Med Pharmacol Sci 2018; 22:7364-70. doi: 10.26355/eurrev_201811_16274 [Crossref] [ Google Scholar]
- Hughes JM, Legnini I, Salvatori B, Masciarelli S, Marchioni M, Fazi F. C/EBPα-p30 protein induces expression of the oncogenic long non-coding RNA UCA1 in acute myeloid leukemia. Oncotarget 2015; 6:18534-44. doi: 10.18632/oncotarget.4069 [Crossref] [ Google Scholar]
- Sun MD, Zheng YQ, Wang LP, Zhao HT, Yang S. Long noncoding RNA UCA1 promotes cell proliferation, migration and invasion of human leukemia cells via sponging miR-126. Eur Rev Med Pharmacol Sci 2018; 22:2233-45. doi: 10.26355/eurrev_201804_14809 [Crossref] [ Google Scholar]
- Chen L, Wang W, Cao L, Li Z, Wang X. Long non-coding RNA CCAT1 acts as a competing endogenous RNA to regulate cell growth and differentiation in acute myeloid leukemia. Mol Cells 2016; 39:330-6. doi: 10.14348/molcells.2016.2308 [Crossref] [ Google Scholar]
- Wang SL, Huang Y, Su R, Yu YY. Silencing long non-coding RNA HOTAIR exerts anti-oncogenic effect on human acute myeloid leukemia via demethylation of HOXA5 by inhibiting Dnmt3b. Cancer Cell Int 2019; 19:114. doi: 10.1186/s12935-019-0808-z [Crossref] [ Google Scholar]
- Wang YH, Lin CC, Hsu CL, Hung SY, Yao CY, Lee SH. Distinct clinical and biological characteristics of acute myeloid leukemia with higher expression of long noncoding RNA KIAA0125. Ann Hematol 2021; 100:487-98. doi: 10.1007/s00277-020-04358-y [Crossref] [ Google Scholar]
- Grasedieck S, Ruess C, Krowiorz K, Lux S, Pochert N, Schwarzer A. The long non-coding RNA cancer susceptibility 15 (CASC15) is induced by isocitrate dehydrogenase (IDH) mutations and maintains an immature phenotype in adult acute myeloid leukemia. Haematologica 2020; 105:e448-53. doi: 10.3324/haematol.2019.235291 [Crossref] [ Google Scholar]
- Wang X, Hou Y, Lyu Y, Zhou J, Zhang X, Hassani MA. LncRNA IRAIN overcomes imatinib resistance in chronic myeloid leukemia via NF-κB/CD44 pathway inhibition. iScience 2024; 27:109851. doi: 10.1016/j.isci.2024.109851 [Crossref] [ Google Scholar]
- Zia S, Rehman N, Ejaz S, Shahid M, Ali M, Shahid R. Transcriptomic and in silico analysis of BLACE (B-cell acute lymphoblastic leukemia expressed), a new non-coding RNA, as a diagnostic biomarker in B-cell ALL. Int J Biochem Cell Biol 2024; 177:106698. doi: 10.1016/j.biocel.2024.106698 [Crossref] [ Google Scholar]
- Pan L, Zheng Y, Zheng H. Long non-coding RNA RP11-252C151 is a potential biomarker of prognosis and hallmark for leukemogenesis in children with B-cell precursor acute lymphoblastic leukemia. Acta Haematol 2024; 147:646-60. doi: 10.1159/000535461 [Crossref] [ Google Scholar]
- Cao L, Hu SY, Pan J, Wang Y, He HL, Lu J. [Expression and clinical significance of long non-coding RNA AC0024541 in children with acute leukemia]. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2020; 28:1433-9. doi: 10.19746/j.cnki.issn.1009-2137.2020.05.001 [Crossref] [ Google Scholar]
- Xie W, Qian TT, Pang Y, Ye X, Huang WH, Zhu P. [Effect of long non-coding RNA MBNL1-AS1 expression on prognosis of acute myeloid leukemia patients]. Zhonghua Yi Xue Za Zhi 2021; 101:934-8. doi: 10.3760/cma.j.cn112137-20200930-02754 [Crossref] [ Google Scholar]
- Xu F, Wu H, Xiong J, Peng T. Long non-coding RNA DLEU2L targets miR-210-3p to suppress gemcitabine resistance in pancreatic cancer cells via BRCA2 regulatio. Front Mol Biosci 2021; 8:645365. doi: 10.3389/fmolb.2021.645365 [Crossref] [ Google Scholar]
- Wang LQ, Wong KY, Li ZH, Chim CS. Epigenetic silencing of tumor suppressor long non-coding RNA BM742401 in chronic lymphocytic leukemia. Oncotarget 2016; 7:82400-10. doi: 10.18632/oncotarget.12252 [Crossref] [ Google Scholar]
- Ghafouri-Fard S, Dashti S, Farsi M, Taheri M. Deleted in lymphocytic leukemia 2 (DLEU2): an lncRNA with dissimilar roles in different cancers. Biomed Pharmacother 2021; 133:111093. doi: 10.1016/j.biopha.2020.111093 [Crossref] [ Google Scholar]
- Tosic N, Tomic Vujovic K, Vukovic V, Kotur N, Stankovic B, Marjanovic I. High expression levels of the long non-coding RNAs lnc-IRF2-3 and lnc-KIAA1755-4 are markers of poor prognosis in chronic lymphocytic leukemia. Int J Mol Sci 2025; 26:1153. doi: 10.3390/ijms26031153 [Crossref] [ Google Scholar]
- Mangiavacchi A, Sorci M, Masciarelli S, Larivera S, Legnini I, Iosue I. The miR-223 host non-coding transcript linc-223 induces IRF4 expression in acute myeloid leukemia by acting as a competing endogenous RNA. Oncotarget 2016; 7:60155-68. doi: 10.18632/oncotarget.11165 [Crossref] [ Google Scholar]
- Fisher L. Retraction: long non-coding RNA KCNQ1OT1 regulates cell proliferation, apoptosis and chemo-sensitivity through modulating the miR-186-5p/NCAM1 axis in acute myeloid leukemia cells. RSC Adv 2021; 11:4616. doi: 10.1039/d1ra90012a [Crossref] [ Google Scholar]
- Dai J, Wang K, Liu T, Wang Q, Pang Y. Retracted Article: Long non-coding RNA KCNQ1OT1 regulates cell proliferation, apoptosis and chemo-sensitivity through modulating the miR-186-5p/NCAM1 axis in acute myeloid leukemia cells. RSC Adv 2019; 9:36256-65. doi: 10.1039/c9ra06378a [Crossref] [ Google Scholar]
- Liccardo F, Iaiza A, Śniegocka M, Masciarelli S, Fazi F. Circular RNAs activity in the leukemic bone marrow microenvironment. Noncoding RNA 2022; 8:50. doi: 10.3390/ncrna8040050 [Crossref] [ Google Scholar]
- Issah MA, Wu D, Zhang F, Zheng W, Liu Y, Fu H. Epigenetic modifications in acute myeloid leukemia: the emerging role of circular RNAs (review). Int J Oncol 2021; 59:107. doi: 10.3892/ijo.2021.5287 [Crossref] [ Google Scholar]
- Jamal M, Song T, Chen B, Faisal M, Hong Z, Xie T. Recent progress on circular RNA research in acute myeloid leukemia. Front Oncol 2019; 9:1108. doi: 10.3389/fonc.2019.01108 [Crossref] [ Google Scholar]
- Saultz JN, Freud AG, Mundy-Bosse BL. MicroRNA regulation of natural killer cell development and function in leukemia. Mol Immunol 2019; 115:12-20. doi: 10.1016/j.molimm.2018.07.022 [Crossref] [ Google Scholar]
- Wiśnik A, Jarych D, Krawiec K, Strzałka P, Potocka N, Czemerska M. Role of microRNAs in acute myeloid leukemia. Genes (Basel) 2025; 16:446. doi: 10.3390/genes16040446 [Crossref] [ Google Scholar]
- Navabi A, Akbari B, Abdalsamadi M, Naseri S. The role of microRNAs in the development, progression and drug resistance of chronic myeloid leukemia and their potential clinical significance. Life Sci 2022; 296:120437. doi: 10.1016/j.lfs.2022.120437 [Crossref] [ Google Scholar]
- Huang HM, Wei YJ, Wang D, Wen XM. [Mechanism of miR-155 promoting drug resistance in childhood acute lymphoblastic leukemia by regulating Wnt/β-catenin signaling pathway]. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2022; 30:418-24. doi: 10.19746/j.cnki.issn.1009-2137.2022.02.016 [Crossref] [ Google Scholar]
- Li LM, Luo FJ, Song X. MicroRNA-370-3p inhibits cell proliferation and induces chronic myelogenous leukaemia cell apoptosis by suppressing PDLIM1/Wnt/β-catenin signaling. Neoplasma 2020; 67:509-18. doi: 10.4149/neo_2020_190612N506 [Crossref] [ Google Scholar]
- Liu Z, Zhong L, Dan W, Chu X, Liu C, Luo X. miRNA-222-3p enhances the proliferation and suppresses the apoptosis of acute myeloid leukemia cells by targeting Axin2 and modulating the Wnt/β-catenin pathway. Biochem Biophys Res Commun 2022; 620:83-91. doi: 10.1016/j.bbrc.2022.06.054 [Crossref] [ Google Scholar]
- Xie Y, Tan L, Wu K, Li D, Li C. miR-26b-5p affects the progression of acute myeloid leukemia by regulating the USP48-mediated Wnt/β-catenin pathway. Crit Rev Eukaryot Gene Expr 2024; 34:33-44. doi: 10.1615/CritRevEukaryotGeneExpr.2024049380 [Crossref] [ Google Scholar]
- Cen HX, Cai SM, Jiang HY, Liao ZM, Han DG. [The mechanism of miR-1294 targeting SOX15 to regulate Wnt/β-catenin signaling pathway and promote the proliferation of acute lymphoblastic leukemia cells in children]. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2023; 31:344-51. doi: 10.19746/j.cnki.issn.1009-2137.2023.02.006 [Crossref] [ Google Scholar]
- Emmrich S, Rasche M, Schöning J, Reimer C, Keihani S, Maroz A. miR-99a/100~125b tricistrons regulate hematopoietic stem and progenitor cell homeostasis by shifting the balance between TGFβ and Wnt signaling. Genes Dev 2014; 28:858-74. doi: 10.1101/gad.233791.113 [Crossref] [ Google Scholar]
- Li C, Gao Q, Wang M, Xin H. LncRNA SNHG1 contributes to the regulation of acute myeloid leukemia cell growth by modulating miR-489-3p/SOX12/Wnt/β-catenin signaling. J Cell Physiol 2021; 236:653-63. doi: 10.1002/jcp.29892 [Crossref] [ Google Scholar]
- Ma L, Wang YY, Jiang P. LncRNA LINC00909 promotes cell proliferation and metastasis in pediatric acute myeloid leukemia via miR-625-mediated modulation of Wnt/β-catenin signaling. Biochem Biophys Res Commun 2020; 527:654-61. doi: 10.1016/j.bbrc.2020.05.001 [Crossref] [ Google Scholar]
- Zhai H, Zhao J, Pu J, Zhao P, Wei J. LncRNA-DUXAP8 regulation of the Wnt/β-catenin signaling pathway to inhibit glycolysis and induced apoptosis in acute myeloid leukemia. Turk J Haematol 2021; 38:264-72. doi: 10.4274/tjh.galenos.2021.2020.0769 [Crossref] [ Google Scholar]
- Chen L, Hu N, Wang C, Zhao H. HOTAIRM1 knockdown enhances cytarabine-induced cytotoxicity by suppression of glycolysis through the Wnt/β-catenin/PFKP pathway in acute myeloid leukemia cells. Arch Biochem Biophys 2020; 680:108244. doi: 10.1016/j.abb.2019.108244 [Crossref] [ Google Scholar]
-
Luo H, Zhu G, Xu J, Lai Q, Yan B, Guo Y, et al. HOTTIP lncRNA promotes hematopoietic stem cell self-renewal leading to AML-like disease in mice. Cancer Cell 2019; 36: 645-59.e8. doi: 10.1016/j.ccell.2019.10.011.
- Du X, Liu H, Yang C, Shi X, Cao L, Zhao X. LncRNA landscape analysis identified LncRNA LEF-AS1 as an oncogene that upregulates LEF1 and promotes survival in chronic lymphocytic leukemia. Leuk Res 2021; 110:106706. doi: 10.1016/j.leukres.2021.106706 [Crossref] [ Google Scholar]
- Chen JJ, Lei P, Zhou M. hsa_circ_0121582 inhibits leukemia growth by dampening Wnt/β-catenin signaling. Clin Transl Oncol 2020; 22:2293-302. doi: 10.1007/s12094-020-02377-9 [Crossref] [ Google Scholar]
- Xia L, Wu L, Bao J, Li Q, Chen X, Xia H. Circular RNA circ-CBFB promotes proliferation and inhibits apoptosis in chronic lymphocytic leukemia through regulating miR-607/FZD3/Wnt/β-catenin pathway. Biochem Biophys Res Commun 2018; 503:385-90. doi: 10.1016/j.bbrc.2018.06.045 [Crossref] [ Google Scholar]
- Ding J, Zhang X, Xue J, Fang L, Ban C, Song B. CircNPM1 strengthens Adriamycin resistance in acute myeloid leukemia by mediating the miR-345-5p/FZD5 pathway. Cent Eur J Immunol 2021; 46:162-82. doi: 10.5114/ceji.2021.108175 [Crossref] [ Google Scholar]
- Lu Y, Zhong L, Luo X, Liu C, Dan W, Chu X. MiRNA-301b-3p induces proliferation and inhibits apoptosis in AML cells by targeting FOXF2 and regulating Wnt/β-catenin axis. Mol Cell Probes 2022; 63:101805. doi: 10.1016/j.mcp.2022.101805 [Crossref] [ Google Scholar]
- Lyu X, Li J, Yun X, Huang R, Deng X, Wang Y. miR-181a-5p, an inducer of Wnt-signaling, facilitates cell proliferation in acute lymphoblastic leukemia. Oncol Rep 2017; 37:1469-76. doi: 10.3892/or.2017.5425 [Crossref] [ Google Scholar]
- Shi LH, Tian L, Wang YF, Liu JS, Guo MF, Liu W. [Effects of MiR-221-mediated Wnt/β-catenin signaling pathway on biological activity of childhood acute lymphoblastic leukemia cells]. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2019; 27:1367-73. doi: 10.19746/j.cnki.issn.1009-2137.2019.05.003 [Crossref] [ Google Scholar]
- Liu B, Ma H, Liu Q, Xiao Y, Pan S, Zhou H. MiR-29b/Sp1/FUT4 axis modulates the malignancy of leukemia stem cells by regulating fucosylation via Wnt/β-catenin pathway in acute myeloid leukemia. J Exp Clin Cancer Res 2019; 38:200. doi: 10.1186/s13046-019-1179-y [Crossref] [ Google Scholar]
- Li H, Xie C, Lu Y, Chang K, Guan F, Li X. Exosomal miR92a promotes cytarabine resistance in myelodysplastic syndromes by activating Wnt/β-catenin signal pathway. Biomolecules 2022; 12:1448. doi: 10.3390/biom12101448 [Crossref] [ Google Scholar]
- Lou Y, Liu L, Zhan L, Wang X, Fan H. miR-187-5p regulates cell growth and apoptosis in acute lymphoblastic leukemia via DKK2. Oncol Res 2016; 24:89-97. doi: 10.3727/096504016x14597766487753 [Crossref] [ Google Scholar]
- Wang Y, Kong D. MicroRNA-136 promotes lipopolysaccharide-induced ATDC5 cell injury and inflammatory cytokine expression by targeting myeloid cell leukemia 1. J Cell Biochem 2018; 119:9316-26. doi: 10.1002/jcb.27208 [Crossref] [ Google Scholar]
- Li G, Song Y, Zhang Y, Wang H, Xie J. miR-34b targets HSF1 to suppress cell survival in acute myeloid leukemia. Oncol Res 2016; 24:109-16. doi: 10.3727/096504016x14611963142254 [Crossref] [ Google Scholar]
- Kang Y, Zhang S, Cao W, Wan D, Sun L. Knockdown of LncRNA CRNDE suppresses proliferation and P-glycoprotein-mediated multidrug resistance in acute myelocytic leukemia through the Wnt/β-catenin pathway. Biosci Rep 2020; 40:BSR20193450. doi: 10.1042/bsr20193450 [Crossref] [ Google Scholar]
- Cuadros M, Andrades Á, Coira IF, Baliñas C, Rodríguez MI, Álvarez-Pérez JC. Expression of the long non-coding RNA TCL6 is associated with clinical outcome in pediatric B-cell acute lymphoblastic leukemia. Blood Cancer J 2019; 9:93. doi: 10.1038/s41408-019-0258-9 [Crossref] [ Google Scholar]
- Chen PH, Liu AJ, Ho KH, Chiu YT, Anne Lin ZH, Lee YT. microRNA-199a/b-5p enhance imatinib efficacy via repressing WNT2 signaling-mediated protective autophagy in imatinib-resistant chronic myeloid leukemia cells. Chem Biol Interact 2018; 291:144-51. doi: 10.1016/j.cbi.2018.06.006 [Crossref] [ Google Scholar]
- Takam Kamga P, Dal Collo G, Cassaro A, Bazzoni R, Delfino P, Adamo A. Small molecule inhibitors of microenvironmental Wnt/β-catenin signaling enhance the chemosensitivity of acute myeloid leukemia. Cancers (Basel) 2020; 12:2696. doi: 10.3390/cancers12092696 [Crossref] [ Google Scholar]
- Elyamany G, Rizwan H, Akhter A, Aljabry MS, Alotaibi S, Albalawi MAH. "Losing the brakes"-suppressed inhibitors triggering uncontrolled Wnt/ß-catenin signaling may provide a potential therapeutic target in elderly acute myeloid leukemia. Curr Issues Mol Biol 2023; 45:604-13. doi: 10.3390/cimb45010040 [Crossref] [ Google Scholar]
- Wakabayashi R, Hattori Y, Hosogi S, Toda Y, Takata K, Ashihara E. A novel dipeptide type inhibitor of the Wnt/β-catenin pathway suppresses proliferation of acute myelogenous leukemia cells. Biochem Biophys Res Commun 2021; 535:73-9. doi: 10.1016/j.bbrc.2020.12.027 [Crossref] [ Google Scholar]
- Hadate Y, Hattori Y, Toda Y, Hosogi S, Okada S, Hayashi Y. Compound #41 targets acute myelogenous leukemia by inhibiting the Wnt/β-catenin signaling pathway. Anticancer Res 2024; 44:4789-99. doi: 10.21873/anticanres.17305 [Crossref] [ Google Scholar]
- Ai Y, Sakamuru S, Imler G, Xia M, Xue F. Improving the solubility and antileukemia activity of Wnt/β-catenin signaling inhibitors by disrupting molecular planarity. Bioorg Med Chem 2022; 69:116890. doi: 10.1016/j.bmc.2022.116890 [Crossref] [ Google Scholar]
- Xi HM, Lu H, Weng XQ, Sheng Y, Wu J, Li L. Combined application of salinomycin and ATRA induces apoptosis and differentiation of acute myeloid leukemia cells by inhibiting WNT/β-catenin pathway. Anticancer Agents Med Chem 2023; 23:1074-84. doi: 10.2174/1871520623666230110121629 [Crossref] [ Google Scholar]
- Qian Y, Zhang X, Mao S, Wei W, Lin X, Ling Q. ACC010, a novel BRD4 inhibitor, synergized with homoharringtonine in acute myeloid leukemia with FLT3-ITD. Mol Oncol 2023; 17:1402-18. doi: 10.1002/1878-0261.13368 [Crossref] [ Google Scholar]
- Al-Hamaly MA, Cox AH, Haney MG, Zhang W, Arvin EC, Sampathi S, et al. Zebrafish drug screening identifies erlotinib as an inhibitor of Wnt/β-catenin signaling and self-renewal in T-cell acute lymphoblastic leukemia. bioRxiv [Preprint]. August 28, 2023. Available from: https://www.biorxiv.org/content/10.1101/2023.08.28.555200v1.full.
- Al-Hamaly MA, Cox AH, Haney MG, Zhang W, Arvin EC, Sampathi S. Zebrafish drug screening identifies erlotinib as an inhibitor of Wnt/β-catenin signaling and self-renewal in T-cell acute lymphoblastic leukemia. Biomed Pharmacother 2024; 170:116013. doi: 10.1016/j.biopha.2023.116013 [Crossref] [ Google Scholar]
- Mao P, Huang C, Li Y, Zhao Y, Zhou S, Zhao Z. Pharmacological targeting of type phosphodiesterase 4 inhibits the development of acute myeloid leukemia by impairing mitochondrial function through the Wnt/β-catenin pathway. Biomed Pharmacother 2023; 157:114027. doi: 10.1016/j.biopha.2022.114027 [Crossref] [ Google Scholar]
- Soldi R, Halder TG, Sampson S, Vankayalapati H, Weston A, Thode T. The small molecule BC-2059 inhibits wingless/integrated (Wnt)-dependent gene transcription in cancer through disruption of the transducin β-like 1-β-catenin protein complex. J Pharmacol Exp Ther 2021; 378:77-86. doi: 10.1124/jpet.121.000634 [Crossref] [ Google Scholar]
- Toden S, Zumwalt TJ, Goel A. Non-coding RNAs and potential therapeutic targeting in cancer. Biochim Biophys Acta Rev Cancer 2021; 1875:188491. doi: 10.1016/j.bbcan.2020.188491 [Crossref] [ Google Scholar]
- Nemeth K, Bayraktar R, Ferracin M, Calin GA. Non-coding RNAs in disease: from mechanisms to therapeutics. Nat Rev Genet 2024; 25:211-32. doi: 10.1038/s41576-023-00662-1 [Crossref] [ Google Scholar]
- Nappi F. Non-coding RNA-targeted therapy: a state-of-the-art review. Int J Mol Sci 2024; 25:3630. doi: 10.3390/ijms25073630 [Crossref] [ Google Scholar]
- Dowdy SF. Endosomal escape of RNA therapeutics: how do we solve this rate-limiting problem?. RNA 2023; 29:396-401. doi: 10.1261/rna.079507.122 [Crossref] [ Google Scholar]
- Baptista B, Riscado M, Queiroz JA, Pichon C, Sousa F. Non-coding RNAs: Emerging from the discovery to therapeutic applications. Biochem Pharmacol 2021; 189:114469. doi: 10.1016/j.bcp.2021.114469 [Crossref] [ Google Scholar]
- Winkle M, El-Daly SM, Fabbri M, Calin GA. Noncoding RNA therapeutics - challenges and potential solutions. Nat Rev Drug Discov 2021; 20:629-51. doi: 10.1038/s41573-021-00219-z [Crossref] [ Google Scholar]
- Vilaça A, de Windt LJ, Fernandes H, Ferreira L. Strategies and challenges for non-viral delivery of non-coding RNAs to the heart. Trends Mol Med 2023; 29:70-91. doi: 10.1016/j.molmed.2022.10.002 [Crossref] [ Google Scholar]
- Kara G, Calin GA, Ozpolat B. RNAi-based therapeutics and tumor targeted delivery in cancer. Adv Drug Deliv Rev 2022; 182:114113. doi: 10.1016/j.addr.2022.114113 [Crossref] [ Google Scholar]
- Li X, Qi H, Cui W, Wang Z, Fu X, Li T. Recent advances in targeted delivery of non-coding RNA-based therapeutics for atherosclerosis. Mol Ther 2022; 30:3118-32. doi: 10.1016/j.ymthe.2022.07.018 [Crossref] [ Google Scholar]
- Yuan Y, Sun W, Xie J, Zhang Z, Luo J, Han X. RNA nanotherapeutics for hepatocellular carcinoma treatment. Theranostics 2025; 15:965-92. doi: 10.7150/thno.102964 [Crossref] [ Google Scholar]
- Witten J, Hu Y, Langer R, Anderson DG. Recent advances in nanoparticulate RNA delivery systems. Proc Natl Acad Sci U S A 2024; 121:e2307798120. doi: 10.1073/pnas.2307798120 [Crossref] [ Google Scholar]
- Garbo S, Maione R, Tripodi M, Battistelli C. Next RNA therapeutics: the mine of non-coding. Int J Mol Sci 2022; 23:7471. doi: 10.3390/ijms23137471 [Crossref] [ Google Scholar]
- Manna I, Fortunato F, De Benedittis S, Sammarra I, Bertoli G, Labate A. Non-coding RNAs: new biomarkers and therapeutic targets for temporal lobe epilepsy. Int J Mol Sci 2022; 23:3063. doi: 10.3390/ijms23063063 [Crossref] [ Google Scholar]
- Karthik S, Mohan S, Magesh I, Bharathy A, Kolipaka R, Ganesamoorthi S. Chitosan nanocarriers for non-coding RNA therapeutics: a review. Int J Biol Macromol 2024; 263:130361. doi: 10.1016/j.ijbiomac.2024.130361 [Crossref] [ Google Scholar]