Bioimpacts. 15:32639.
doi: 10.34172/bi.32639
Original Article
Resolvin D1 alleviates lipopolysaccharide-induced in vitro corneal inflammation by modulating macrophage polarization through upregulation of c-Fos expression
Qin Qin Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Resources, Validation, Visualization, Writing – original draft, 1, 2, 3, # 
Yinru Wang Data curation, Formal analysis, Investigation, Visualization, Writing – original draft, 1, #
Yajun Liu Formal analysis, 1
Weiwei An Formal analysis, Investigation, Visualization, 2, 4
Zifang He Formal analysis, Investigation, 1
Feifei Chen Data curation, Investigation, 1
Xinxuan Cheng Investigation, 1
Deyan Chen Conceptualization, Methodology, Supervision, Validation, Writing – review & editing, 5, *
Zhenggao Xie Conceptualization, Funding acquisition, Project administration, Resources, Supervision, Validation, Writing – review & editing, 1, 2, 3, *
Author information:
1Nanjing Drum Tower Hospital, The Affiliated Hospital of Nanjing University Medical School, Nanjing, Jiangsu, 210008, China
2Nanjing Drum Tower Hospital, Clinical College of Nanjing Medical University, Nanjing, Jiangsu, 210008, China
3Nanjing Drum Tower Hospital, Clinical College of Traditional Chinese Medicine, Nanjing University of Chinese Medicine, Nanjing, Jiangsu, 210008, China
4Yancheng Tinghu District People’s Hospital, Yancheng, Jiangsu, 224005, China
5Key Laboratory of Infection and Immunity of Anhui Higher Education Institutes, Bengbu Medical University, No. 2600, Donghai Avenue, Bengbu, Anhui, 233030, China
#,*These authors contributed equally to this work.
Abstract
Introduction:
Bacterial keratitis refers to a prevalent sight-threatening ophthalmologic infection. Owing to the challenge of antimicrobial resistance in treating bacterial keratitis, novel therapeutic strategies are needed. Resolvin D1 (RvD1), an endogenous lipid mediator, exhibits anti-inflammatory and immune-regulatory effects. The present study was aimed at investigating whether RvD1 alleviates lipopolysaccharide-induced inflammation to protect corneal fibroblasts and explore its potential mechanisms.
Methods:
In this study, Raw264.7 cells were polarized towards M1 or M2 macrophages by the addition of lipopolysaccharides LPS or interleukin (IL)-4, respectively, and were treated with or without RvD1. Flow cytometry and Western blot were used to determine the expression of M1 and M2-related markers. EdU assay and trans-well assay were performed to detect the proliferation and migration ability of corneal fibroblasts. Bioinformatics analysis (GO and KEGG) of RNA-seq was applied to explore the RvD1-related signaling pathways. siRNA-c-Fos was further used to confirm the role of Fos expression in RvD1-mediated macrophage polarization. Flow cytometry and Western blotting analysis were performed to demonstrate that RvD1 alleviated LPS-induced inflammation by suppressing M1 macrophage polarization, facilitating M2 macrophage polarization, and increasing corneal fibroblast proliferation. Bioinformatics analysis identified PI3K-AKT, IL-17, and MAPK signaling pathways as potential targets of RvD1 in corneal inflammation.
Results:
Enrichment analysis indicated that the RvD1 target gene showed a strong relationship to the regulation of macrophage polarization. RvD1 highly upregulated M2 macrophages by promoting c-Fos expression and enhanced the proliferation and migration of mouse corneal fibroblasts through modulating c-Fos expression.
Conclusion:
Our findings reveal that RvD1 conferred protective effects against LPS-induced inflammation by enhancing M2 macrophage polarization through the promotion of c-Fos expression. Thus, RvD1 may be a potential therapeutic compound for enhancing corneal fibroblast proliferation and migration while attenuating inflammation.
Keywords: Corneal inflammation, Macrophage polarization, Resolvin D1
Copyright and License Information
© 2025 The Author(s).
This work is published by BioImpacts as an open access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (
http://creativecommons.org/licenses/by-nc/4.0/). Non-commercial uses of the work are permitted, provided the original work is properly cited.
Funding Statement
This research was supported by the medical research key project of Jiangsu Provincial Health Commission of China (ZD2022022) and the Clinical Trials from the Affiliated Drum Tower Hospital, Medical School of Nanjing University (2021-LCYJ-PY-25, 2023-LCYJ-MS-30) and the New Medical Technology Development Project of Nanjing Drum Tower Hospital (XJSFZLX202330).
Introduction
Bacterial keratitis is a common ocular infection and one of the leading causes of irreversible corneal blindness.1 Its increasing incidence can be attributed to ocular trauma and the excessive use of contact lenses.2 Subsequently, microbial infections may progress to corneal ulceration and perforation, potentially leading to severe vision loss. Meanwhile, antimicrobial resistance is prevalent in bacterial keratitis. Hence, there is an urgent need to explore practical strategies for treating bacterial keratitis.3,4
Macrophages are essential in immune defense, self-stabilization, and surveillance, and exhibit two phenotypes, namely M1 and M2 macrophages.5 M1 macrophages produce cytokines, which include tumor necrosis factor-α (TNF-α), inducible nitric-oxide synthase (iNOS), and CD86, which are crucial for pro-inflammatory effects.6 Conversely, M2 macrophages release cytokines, including interleukin (IL)-10, Arginase 1 (Arg-1), and CD206, which primarily play an anti-inflammatory role and have beneficial effects on keratitis.7,8 M2 macrophages promote wound healing, thereby preventing severe immunopathology.9 Arg-1 and CD206 are mostly engaged in the modulation of the anti-inflammatory response and tissue remodeling in the host, which are the two main polarization directions of activated macrophages.10 Furthermore, the precise regulation and balance between M1 and M2 homeostasis are essential in the modulation of acute inflammation. Therefore, understanding how to regulate macrophage differentiation effectively is of great value for developing a novel therapeutic strategy to manage bacterial keratitis. Resolvin D1 (RvD1), as a specialized pro-resolving mediator produced from docosahexaenoic acid, an omega-3 polyunsaturated fatty acid in the body, exerts anti-inflammatory and immunomodulatory properties.11 These receptors have been widely distributed on the cell surface of various human tissues. RvD1 exhibits its biological actions through the mediation of Orphan GPR32 and ALX/FPR2.12 By reducing the infiltration of inflammatory cells, RvD1 may ultimately achieve anti-inflammatory effects and tissue regeneration, thus hindering the secretion of inflammatory factors and promoting macrophage polarization.13,14 RvD1 is highly expressed in the corneal epithelium and induces macrophage polarization to the M2-type, further promoting corneal inflammation resolution.12,15 Treatment with RvD1 could inhibit the secretion of TNF-α, IL-6, and IL-8 in human corneal epithelial cells.16 By modulating anti-inflammatory cytokines, including IL-10, RvD1 enhanced M2 polarization in LPS-induced bacterial keratitis mouse models. 17 Our study aimed to address these literature gaps.
Using an LPS-induced inflammatory animal model, this work aimed at examining the regulatory function of RvD1 and exploring its mechanism of modulating macrophage polarization. Bioinformatics analysis was performed to predict the potential signaling pathways of RvD1. This study provides valuable insights into the immunopathogenesis of corneal inflammation, and its findings may contribute to the development of novel therapeutic strategies for the management of bacterial keratitis.
Materials and Methods
Reagents
RvD1 (Cat#: 10012554) was supplied by Cayman Chemical (Michigan, USA). IL-4 (Cat#: 200-04 and 214-14) was obtained from PeproTech (New Jersey, USA), and LPS (Cat#: MB5198) was from Meilun Biotechnology Co., Ltd. (Dalian, China).
Cell culture
Murine (mouse) macrophage cell line (Raw264.7) and Tohoku Hospital pediatrics-1 (THP-1) cells were acquired from the Cell Bank, Chinese Academy of Sciences. Mouse corneal fibroblast (MCF) cells were bought from Yuchun Biotechnology Co., Ltd. (Shanghai, China). Dr. He (Jiangsu Provincial People’s Hospital, Nanjing, China) provided the human umbilical vein endothelial cells (HUVECs). Raw264.7 cells were cultivated in Dulbecco’s modified Eagle medium (DMEM) (11995500, Gibco, China), while THP-1 cells were cultivated in 1640 medium (Cat#:11875500, Gibco, China). The DMEM and 1640 medium were added with 10% fetal bovine serum (Cat#: FB25015, CLARK Bioscience, USA). MCFs were maintained in a special culture medium (Cat#: CCR0194, Yuchun Bio) and human umbilical vein endothelial cells (HUVECs) in endothelial cell medium (ECM) (Cat#: 1001, Sciencell). Cells were cultured until 80% confluence at 37°C with 5% CO2.18
Treatment of cultured cells
Raw264.7 cells were subject to treatment with IL-4 (10 ng/mL) or co-treated with RVD1 (100 nM) for 24 hours; at the same time, the cells were treated with LPS (100 n/mL) or RVD1 was co-treated with LPS for 24 hours. Then the cells were collected and the proteins were extracted for subsequent detection. Raw264.7 and THP-1 cells were transfected with siRNA-c-Fos (target sequence: 5’CGAGCAGATGAACTTAACAAT3’) to inhibit c-Fos expression. The transfected Raw264.7 cells were subject to treatment with IL-4 or RvD1 and co-cultured with MCFs to detect cell migration and proliferation. The transfected THP-1 cells were exposed to treatment with IL-4 or RvD1 and co-cultured with HUVECs to detect cell migration and proliferation.
Flow cytometry
Cells were centrifuged (500 × g, 4 °C, 5 min) and rinsed with 2 mL phosphate-buffered saline (PBS) three times. The cell suspension was labeled with the anti-mouse antibodies CD11b-FITC (Cat#: 557396, BD Pharmingen, USA), CD86-PE (Cat#: 560957, BD Pharmingen), and CD206-APC (Cat#: 561763, BD Pharmingen) in the dark (4 °C, 30 min). Before labeling, mouse Fc block (Cat#: 564219, BD Pharmingen) was used for pre-incubation. Flow cytometry (FACSCalibur, BD Biosciences, USA) was employed to analyze the samples.19,20
RNA sequencing
The extraction of total RNA was carried out with the TRIzol reagent (Invitrogen, Carlsbad, USA) and quantified with the NanoDrop ND-1000 spectrophotometer (Wilmington, DE, USA). Denaturing agarose gel electrophoresis was performed to confirm RNA integrity. Poly(A) RNA was fragmented and reverse-transcribed into cDNA (NEB, cat#: e6150, USA). Previously, blunt ends were treated with an A-base to render them set up for adapter ligation. The ligated products were subject to PCR amplification, and the final cDNA library was assembled.21 Ultimately, 2 × 150-bp paired-end reads were sequenced on the Illumina NovaseqTM 6000 platform (LC-Bio Technology Co., Ltd., Hangzhou, China).22
5-Ethynyl-2-deoxyuridine (EdU) assay
Raw264.7 cells were exposed to different treatments (LPS, IL-4, siRNA-c-Fos, or a combination) and co-cultured with MCF cells via the trans-well system. The measurement of cell proliferation was made using the EdU kit (Cat#: MA0425, Meilun Biotech, China) following the guidance of the manufacturer. Images were recorded with a fluorescence microscope (LIOO, China). The proliferation rate was calculated as the ratio of the number of EdU-positive cells to that of the total cells (determined by DAPI staining).
Immunofluorescence analysis
Cells were treated with 4% paraformaldehyde, blocked with normal goat serum (Cat#: WGAR1009-5, Servivebio, China), and incubated with anti-c-FOS antibodies (1:100 dilution) at 4°C overnight. In addition, FITC-conjugated secondary antibodies were adopted for immunolabeling detection. Nuclei were stained with DAPI. Immunofluorescence was visualized using Olympus Fluoview FV3000.23,24
Migration assay
Trans-well chambers (Corning Costar Corporation, Corning, NY, USA) were used for assessing the changes in MCF migration following co-cultivation with Raw264.7 cells. MCF cells were cultivated in the upper chamber, whereas drug-treated Raw264.7 cells were cultivated in the lower chamber to induce MCF migration. Then, cells were stained with a 0.1% crystal violet solution, and cell counting was performed under an inverted microscope across five fields.
Western blotting analysis
Extracted proteins were separated using 10% SDS–PAGE and transferred onto polyvinylidene fluoride (PVDF) membranes (Millipore, Germany). The membranes were exposed to incubation with the primary and secondary antibodies. The antibodies used were as follows: CD206 (ab64693, Abcam), Arg-1 (Cat#:93668S, CST, USA), CD163 (Cat#: ab182422, Abcam, USA), c-Fos (Cat#:sc-8047, Santa Cruz, USA), and anti-β-actin (Cat#:4970S, CST, USA). Bands were visualized through an electrochemiluminescence (ECL) detection system, TANON 5200 (Tanon, Shanghai, China).25,26
Bioinformatics analysis
Enrichment analysis of GO terms and KEGG pathways of key target proteins was conducted using the DAVID database, and those with *P < 0.05 were thought to be of significance. Differentially expressed genes (DEGs) are represented by the heat map. Results were analyzed visually.
Statistical analysis
Results are shown to be the mean ± standard error of the mean (SEM). Statistical significance was defined at *P < 0.05. Data were explored with one-way ANOVA with the Student–Newman–Keuls test in SPSS version 20 (SPSS, Inc., USA).
Results
RvD1 inhibits M1 macrophage polarization but stimulates M2 macrophage polarization in vitro
Promoting the M1-to-M2 transition of macrophages represents a highly effective approach to mitigating inflammation.27 Once tissue injury occurs, M2 macrophages are involved in tissue repair and reconstruction, thus reducing the inflammatory response. It was indicated that RvD1 reduced the proportion of M1 macrophages in LPS-treated Raw 267.4 cells (Fig. 1A) and increased that of M2 macrophages, thereby promoting M2 macrophage polarization (Fig. 1B). Moreover, in the IL-4 group, following RvD1 treatment, M2 markers CD206, Arg-1, and CD163 were upregulated (Fig. 1C). These findings indicate that RvD1 facilitated the transformation of inflammatory (M1) macrophages to anti-inflammatory (M2) macrophages, thus mitigating the inflammation triggered by LPS.
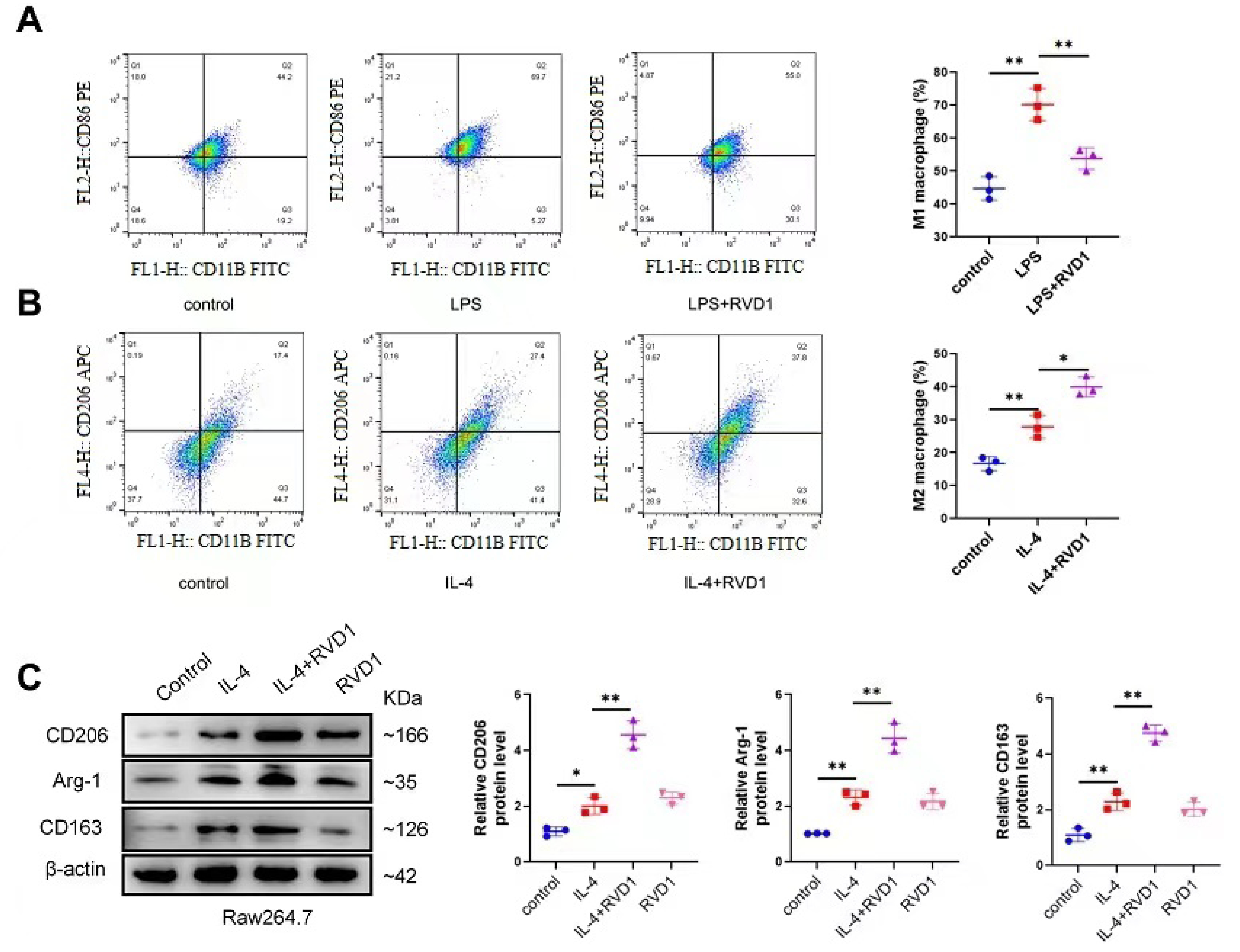
Fig. 1.
RvD1 inhibits M1 macrophages and promotes M2 macrophage polarization.Percentages of M1-like macrophages (A) (CD11b+, CD86+) and M2-like macrophages (B) (CD11b+, CD206+) were analyzed by flow cytometry. (C) Protein levels of M2 macrophage markers CD206, Arg-1, and CD163 were determined by Western blot. *P < 0.05, **P < 0.01.
.
RvD1 inhibits M1 macrophages and promotes M2 macrophage polarization.Percentages of M1-like macrophages (A) (CD11b+, CD86+) and M2-like macrophages (B) (CD11b+, CD206+) were analyzed by flow cytometry. (C) Protein levels of M2 macrophage markers CD206, Arg-1, and CD163 were determined by Western blot. *P < 0.05, **P < 0.01.
RvD1 facilitates the proliferation and migration of corneal fibroblasts by activating M2 macrophages in vitro
To examine the impact of RvD1 in M2 macrophages on the proliferation and migration of MCFs, we co-cultured mouse corneal fibroblasts with RvD1-treated or untreated M2 macrophages (IL-4 induced). Subsequently, EdU assays were performed to detect the fold changes in mouse corneal fibroblasts. Only IL-4-induced M2 macrophages promoted the proliferation of corneal fibroblasts. Additionally, treatment with RvD1 further enhanced the proliferation of corneal fibroblasts (Fig. 2A) and their migration (Fig. 2B). The obtained findings indicate that RvD1 enhanced the polarisation into M2 macrophages (IL-4 induction) and facilitated the migration of corneal fibroblasts, thereby enhancing corneal wound healing in vitro.
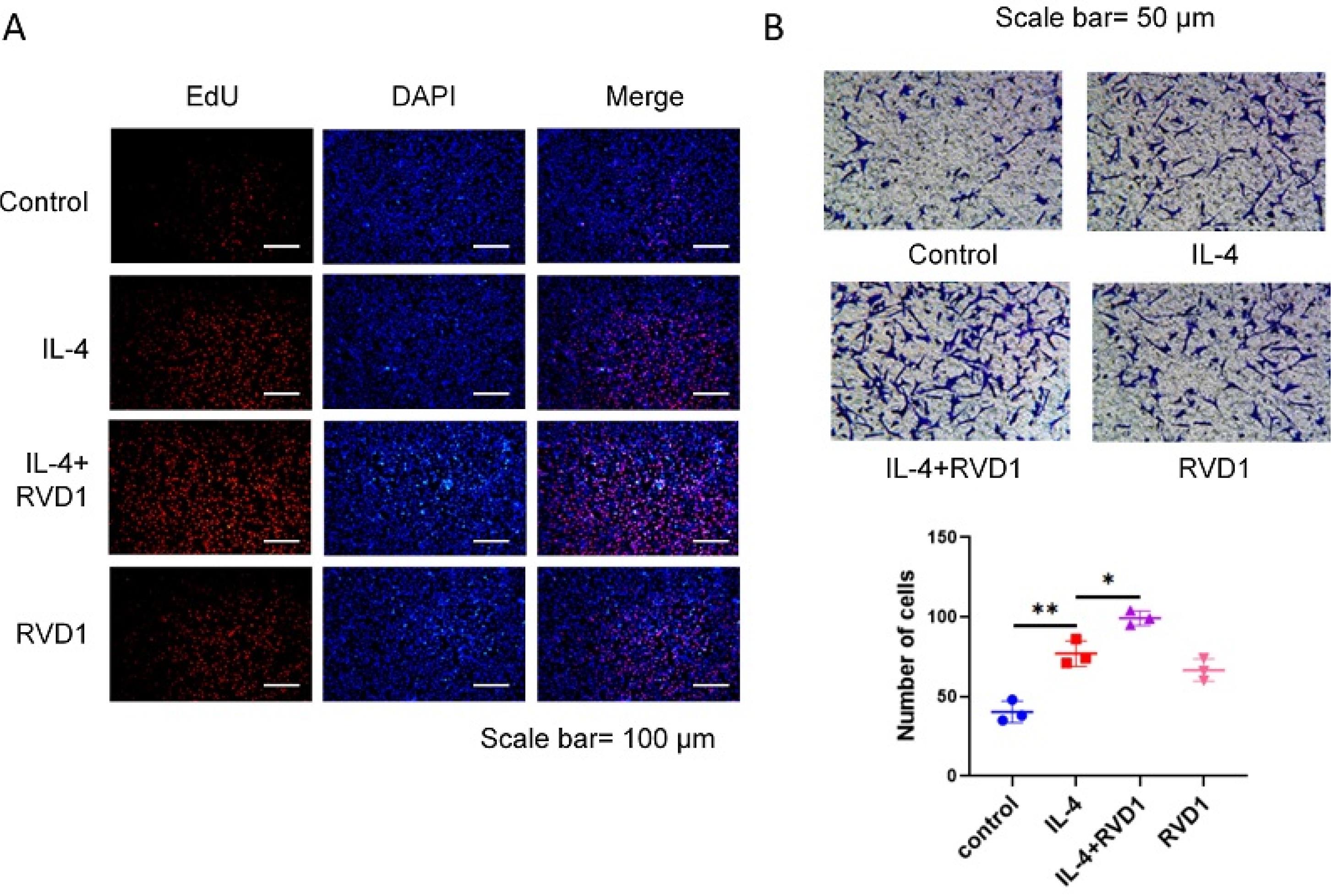
Fig. 2.
RvD1 promotes corneal fibroblast proliferation and migration by regulating M2 macrophages. (A) After corneal fibroblasts and macrophages were co-cultured for 24 hours, the EdU assay was used to detect the corneal fibroblast proliferation. (B) The migration ability of corneal fibroblasts was tested using the trans-well assay (*P < 0.05, **P < 0.01).
.
RvD1 promotes corneal fibroblast proliferation and migration by regulating M2 macrophages. (A) After corneal fibroblasts and macrophages were co-cultured for 24 hours, the EdU assay was used to detect the corneal fibroblast proliferation. (B) The migration ability of corneal fibroblasts was tested using the trans-well assay (*P < 0.05, **P < 0.01).
RvD1 regulates M2 macrophage polarization
RNA sequencing was performed on IL-4-induced M2 macrophages with or without RvD1 treatment to investigate the potential target genes of RvD1-mediated M2 macrophage polarization. The volcano plot and heat map show that the c- Fos gene in the RvD1-treated + IL-4 group was upregulated relative to the PBS-treated + IL-4 group (Fig. 3A and 3B). KEGG enrichment analysis showed that RvD1 treatment regulates the PI3K-AKT, IL-17, and MAPK signaling pathways (Fig. 3C). These three signaling pathways are strongly associated with the M1/M2 polarization. Furthermore, GO enrichment analysis revealed that in the biological process category, RvD1 treatment is engaged in positive regulation of extracellular signal-regulated kinases 1 and 2 (ERK1/2), nuclear factor kappa B (NF-κB) signaling pathway, and cell migration; in the cellular component category, it is related to endoplasmic reticulum stress and extracellular matrix; and in the molecular function category, it relates to protein binding, molecular function, and DNA binding (Fig. 3D). The original sequencing data of the downcomer were in FASTQ format. Quality control was performed using fastp (https://github.com/OpenGene/fastp) with default parameters, including removal of adapters, duplicate sequences, and low-quality reads. The cleaned sequencing data were then aligned to the human reference genome (Homo sapiens, GRCh38) using HISAT2 (https://ccb.jhu.edu/software/hisat2), and the alignment results were obtained in BAM format.
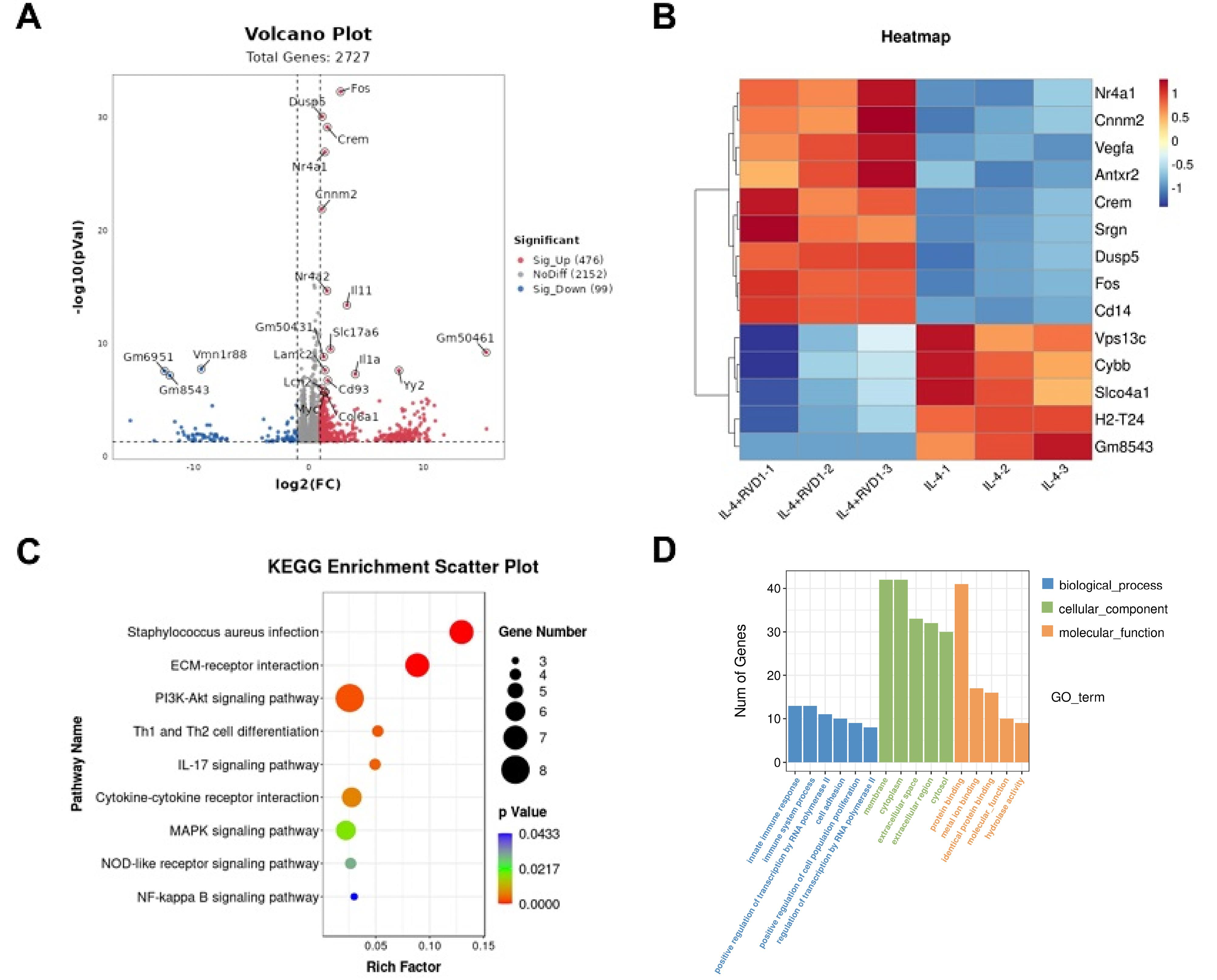
Fig. 3.
RvD1 regulates the gene expression profile of M2 macrophages induced by IL-4. (A) Volcano plot showed all genes; (B) Heat map showed DEGs; (C) Scatter plot showed KEGG pathway enrichment analysis; (D) Histogram used for GO analysis. DEG, differentially expressed gene; GO, gene ontology; RvD1, Resolvin D1; KEGG, Kyoto Encyclopedia of Genes and Genomes.
.
RvD1 regulates the gene expression profile of M2 macrophages induced by IL-4. (A) Volcano plot showed all genes; (B) Heat map showed DEGs; (C) Scatter plot showed KEGG pathway enrichment analysis; (D) Histogram used for GO analysis. DEG, differentially expressed gene; GO, gene ontology; RvD1, Resolvin D1; KEGG, Kyoto Encyclopedia of Genes and Genomes.
RvD1 enhances M2 macrophage polarization by promoting c- Fos expression
The RNA sequencing results suggest that RvD1 increased the mRNA levels of c-Fos in IL-4-stimulated M2 macrophages. The RvD1 + IL-4 group exhibited a notable elevation in c-Fos protein levels compared to the PBS + IL-4 group. Additionally, siRNA-c-Fos treatment abolished the increased c-Fos protein levels induced by RvD1 treatment (Fig. 4A). Elevated c-Fos expression was stimulated by IL-4 + RvD1 treatment, and the levels of M2 macrophage markers, namely Arg-1 and CD163, were significantly increased. In contrast, in the IL-4 + RvD1 + siRNA-c-Fos group, the levels of Arg-1 and CD163 were markedly reduced following the decrease in c-Fos expression (Fig. 4B). The findings indicate that RvD1 enhanced M2 macrophage polarization through regulating c-Fos expression.
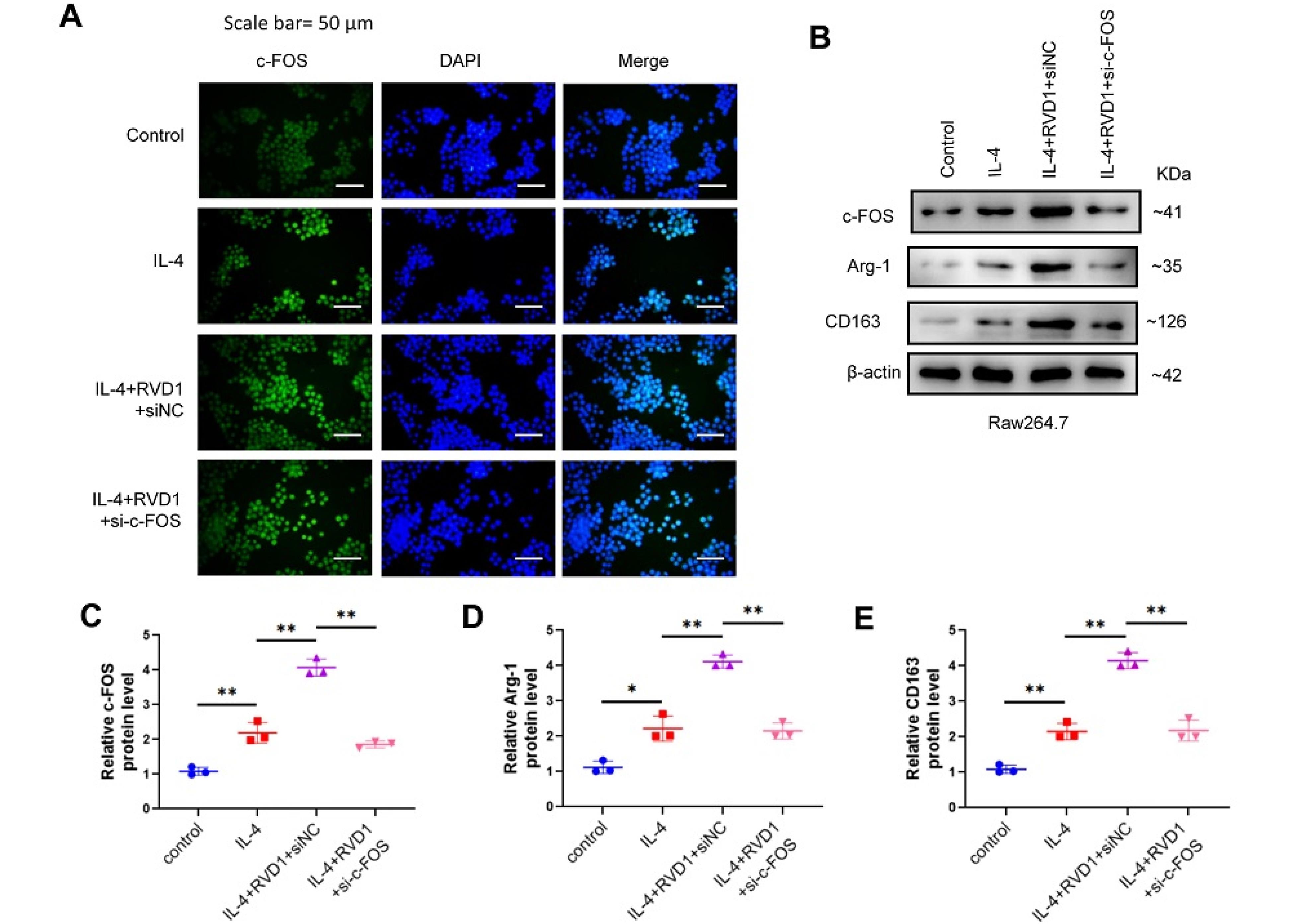
Fig. 4.
RvD-1 enhances M2 macrophage polarization through the c-Fos gene.After Raw264.7 cells were interfered with siRNA of the c-Fos gene, (A) the protein level of c-Fos was analyzed by cell immunofluorescence experiment; (B) the levels of c-Fos, Arg-1, and CD163 were detected by western blot (*P < 0.05, **P < 0.01).
.
RvD-1 enhances M2 macrophage polarization through the c-Fos gene.After Raw264.7 cells were interfered with siRNA of the c-Fos gene, (A) the protein level of c-Fos was analyzed by cell immunofluorescence experiment; (B) the levels of c-Fos, Arg-1, and CD163 were detected by western blot (*P < 0.05, **P < 0.01).
M2-type macrophages induced by RvD1 enhance the proliferation and migration of mouse corneal fibroblasts by modulating c- Fos expression
The results of the EdU assays indicated that a decrease in the protein level of c-Fos significantly abolished the positive regulation of RvD1 on M2 macrophages (Fig. 5A). This sheds light on the intricate relationship between c-Fos in RvD1-induced macrophages and their influence on the behavior of MCFs. The findings of the trans-well assays were consistent with those of the EdU assays (Fig. 5B). Therefore, siRNA-c-Fos transfected into macrophages can significantly inhibit the impact of RvD1-induced M2-type macrophages on enhancing the migration of MCFs.
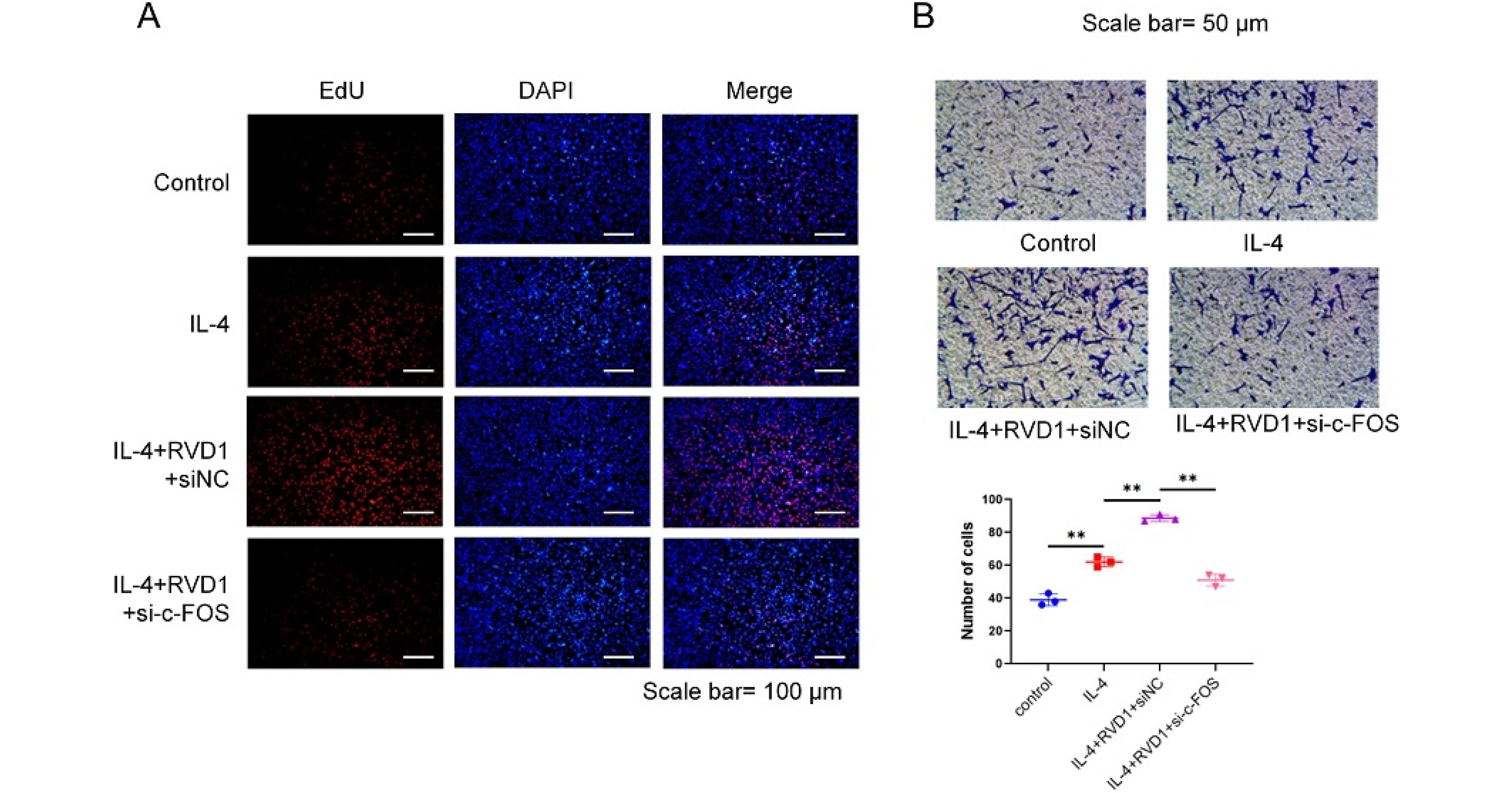
Fig. 5.
RvD1 regulates the proliferation and migration of corneal fibroblasts dependent on macrophage c-Fos gene expression. In vitro experiments, after interfering c-Fos gene of Raw264.7 cells with siRNA, they were co-cultured with corneal fibroblasts for 24 h. (A) The proliferation ability of corneal fibroblasts was tested using EdU assay; (B) the migration ability of corneal fibroblasts was detected by trans-well assay (*P < 0.05, **P < 0.01).
.
RvD1 regulates the proliferation and migration of corneal fibroblasts dependent on macrophage c-Fos gene expression. In vitro experiments, after interfering c-Fos gene of Raw264.7 cells with siRNA, they were co-cultured with corneal fibroblasts for 24 h. (A) The proliferation ability of corneal fibroblasts was tested using EdU assay; (B) the migration ability of corneal fibroblasts was detected by trans-well assay (*P < 0.05, **P < 0.01).
Discussion
Currently, there is increasing resistance to commonly prescribed antibiotics in treating bacterial keratitis.28 This study revealed that, in an LPS-induced mouse model, the endogenous lipid mediator RvD1 plays a protective role by triggering M2 macrophage polarization via the upregulation of c-Fos.
RvD1 significantly hindered the secretion of pro-inflammatory cytokines, which included those of TNF-α, interferon-gamma (IFN-γ), IL-1β, C-X-C motif chemokine ligand 1 (CXCL1), and S100A8/9.29 In various infection models, such as bacterial and fungal keratitis, these cytokines enhance the recruitment of neutrophils in the corneal stroma.30 By suppressing the activation of the NF-κB signaling pathway, RvD1 can reduce the secretion of pro-inflammatory cytokines.16,31,32 Most importantly, RvD1 can rapidly halt neutrophil migration and reduce the inflammatory responses in infectious keratitis.33 RvD1 could lessen the severity of Pseudomonas keratitis or prevent the development of Pseudomonas keratitis by reducing neutrophil recruitment, promoting M2 macrophage polarization, and inhibiting inflammatory responses.29 Neutrophils are the primary effectors of acute inflammatory processes, whereas macrophages are essential in regulating the onset and resolution of inflammation following bacterial infections.
In corneal inflammation, M1 macrophages exhibit anti-bacterial ability, but they lead to corneal clarity loss and oxidative tissue damage.34 Conversely, M2 macrophages repair tissue by reducing the severity of corneal inflammatory lesions and removing necrotic cell residues.35-37 Furthermore, RvD1 treatment modifies several immune processes involved in the pathogenesis of bacterial keratitis, including neutrophil infiltration, secretion of pro- and anti-inflammatory cytokines, and macrophage polarization.30,38,39 RvD1 promotes M2 macrophage polarization via multiple pathways, such as G protein-coupled receptor 32 (GRP32), AGE, and NF/κB pathway.17,40-43 Our study revealed that RvD1 increased the expression of levels of IL-4-induced CD206, Arg-1, and CD163 proteins. According to the obtained findings, RvD1 alleviated the inflammatory response by modulating the M1-to-M2 transition, thus exerting anti-inflammatory effects in an LPS-induced model, suggesting that RvD1 is a promising target for treating bacterial keratitis.
In various diseases, M2 macrophages promote tissue healing. For example, M2-derived exosomal miR-590-3p decreased the inflammatory signals and stimulated the production of epithelial cells.44 The polarization of M2 macrophages could also promote wound repair in diabetic patients.45-47 Increasing evidence has shown that M2 macrophages exert a beneficial impact on the prevention of kidney injury and vascular and osteogenic repair.48-54 Moreover, M2 macrophages possess anti-inflammatory effects and result in the restoration of the ocular surface in dry eyes.55 The proliferation and migration of corneal fibroblasts contribute to corneal epithelial wound healing.56 The findings conform to ours, which showed that M2 macrophages (induced by IL-4) promote the proliferation of corneal fibroblasts, and RvD1 increased the proliferation and migration of corneal stromal fibroblasts through regulating the M2 macrophage polarization. Therefore, our results suggest that RvD1 not only inhibits bacteria-induced corneal inflammation by driving the M2-type polarization but also enhances the proliferation and migration of corneal fibroblasts, thereby promoting the repair of ulcerated corneas.
C-Fos is crucial for controlling gene expression in the nucleus, including the transacting serum response factor (SRF), the ternary complex factor -Elk-1 (TCF/Elk-1), the cAMP-response element binding protein (CREB), protein kinase A and C (PKA, PKC), calcium-calmodulin dependent kinases (CaM kinase), and mitogen-activated kinases (MAP-kinase).57 Several studies have shown that c-Fos and c-Jun enhance the transcription and translation of macrophage-specific functional genes by increasing their expression as monocytes differentiate into macrophages.58
The inhibition of the Fos/MAPK pathway hinders M1 macrophage polarization and facilitates the conversion of M1 to M2 macrophages. This transition results in a decrease in the production of cytokines and alleviation of atherosclerosis.59 Currently, it is understood that c-Fos controls the secretion of inflammatory cytokines.60-62 Pneumonia infection causes the activation of c-Fos in human coronary artery endothelial cells, leading to an accelerated disease course.63 Jin has suggested that Fos/MAPK could hinder M1-macrophage polarization triggered by a high-fat diet, regulate cytokine release, and alleviate atherosclerosis.44 Chen et al. have suggested that c-Fos is critical for macrophage subtype polarization.64 In the present study, c-Fos expression was significantly higher in the RvD1 + IL-4 groups compared to the PBS + IL-4 group, and this RvD1-induced increase in c-Fos expression was abolished by siRNA-c-Fos treatment. Furthermore, downregulation of c-Fos protein levels—whether mediated by IL-4, RvD1, or siRNA-c-Fos—markedly reduced the expression of Arg-1 and CD163.
The study has some limitations. The first is whether RvD1 has any toxic side effects. The second is that our study was not conducted on human beings due to ethical limitations. We will further improve it in the future.
Conclusion
In conclusion, this study is the first to demonstrate that RvD1 promotes M2-type macrophage polarization by regulating c-Fos expression, thereby influencing the proliferation and migration of mouse corneal fibroblasts. RvD1 may serve as a protective agent against bacterial keratitis in mice. Future studies will further investigate its anti-inflammatory and anti-infective effects in human bacterial keratitis.
Research Highlights
What is the current knowledge?
-
Bacterial keratitis is a sight-threatening infection with prevalent antimicrobial resistance.
-
Macrophages have M1 (pro-inflammatory) and M2 (anti-inflammatory) phenotypes.
-
RvD1 has anti-inflammatory effects and can promote M2 macrophage polarization.
-
M2 macrophages aid tissue repair and corneal inflammation resolution.
-
RvD1 acts via GPR32/ALX/FPR2 receptors to regulate immune responses.
What is new here?
-
RvD1 alleviates corneal inflammation by upregulating c-Fos to enhance M2 polarization.
-
RvD1 targets PI3K-AKT, IL-17, and MAPK pathways in corneal inflammation.
-
c-Fos is key for RvD1-mediated M2 polarization and corneal fibroblast function.
-
SiRNA-c-Fos abolishes RvD1’s effects on M2 polarization and fibroblast activity.
-
RvD1 boosts corneal fibroblast proliferation/migration via c-Fos-modulated M2.
Competing Interests
The author declares that no conflict of interest.
Ethical Approval
This study was approved by the Laboratory Animal Ethics Committee of Nanjing Hospital Affiliated to Nanjing Medical University (Ethic Code No: DWSY-24113569).
Acknowledgements
The authors would like to thank all the staff of the Laboratory of Drum Tower Hospital, affiliated with Nanjing University, for their valuable contributions to this research. This research was supported by the medical research key project of Jiangsu Provincial Health Commission of China (ZD2022022) and the Clinical Trials from the Affiliated Drum Tower Hospital, Medical School of Nanjing University (2021-LCYJ-PY-25, 2023-LCYJ-MS-30).
References
- Acharya M, Farooqui JH, Jain S, Mathur U. Pearls and paradigms in infective keratitis. Rom J Ophthalmol 2019; 63:119-27. [ Google Scholar]
- Ting DS, Ho CS, Deshmukh R, Said DG, Dua HS. Infectious keratitis: an update on epidemiology, causative microorganisms, risk factors, and antimicrobial resistance. Eye (Lond) 2021; 35:1084-101. doi: 10.1038/s41433-020-01339-3 [Crossref] [ Google Scholar]
- Wu Z, Sun W, Wang C. Clinical characteristics, treatment, and outcomes of pembrolizumab-induced uveitis. Invest New Drugs 2024; 42:510-7. doi: 10.1007/s10637-024-01464-w [Crossref] [ Google Scholar]
- Wu Z, Sun W, He B, Wang C. Clinical characteristics, treatment, and outcomes of nivolumab-induced uveitis. ImmunopharmacolImmunotoxicol 2025; 47:222-7. doi: 10.1080/08923973.2025.2461056 [Crossref] [ Google Scholar]
- Zhou Z, Sui X, Cao Z, Li X, Qing L, Tang J. Substance P promote macrophage M2 polarization to attenuate secondary lymphedema by regulating NF-kB/NLRP3 signaling pathway. Peptides 2023; 168:171045. doi: 10.1016/j.peptides.2023.171045 [Crossref] [ Google Scholar]
- Yunna C, Mengru H, Lei W, Weidong C. Macrophage M1/M2 polarization. Eur J Pharmacol 2020; 877:173090. doi: 10.1016/j.ejphar.2020.173090 [Crossref] [ Google Scholar]
- Ren Y, Khan FA, Pandupuspitasari NS, Zhang S. Immune evasion strategies of pathogens in macrophages: the potential for limiting pathogen transmission. Curr Issues Mol Biol 2017; 21:21-40. [ Google Scholar]
- Shapouri-Moghaddam A, Mohammadian S, Vazini H, Taghadosi M, Esmaeili SA, Mardani F. Macrophage plasticity, polarization, and function in health and disease. J Cell Physiol 2018; 233:6425-40. doi: 10.1002/jcp.26429 [Crossref] [ Google Scholar]
- Arnold L, Henry A, Poron F, Baba-Amer Y, van Rooijen N, Plonquet A. Inflammatory monocytes recruited after skeletal muscle injury switch into antiinflammatory macrophages to support myogenesis. J Exp Med 2007; 204:1057-69. doi: 10.1084/jem.20070075 [Crossref] [ Google Scholar]
- Liu YC, Zou XB, Chai YF, Yao YM. Macrophage polarization in inflammatory diseases. Int J Biol Sci 2014; 10:520-9. doi: 10.7150/ijbs.8879 [Crossref] [ Google Scholar]
- Roohbakhsh A, Etemad L, Karimi G. Resolvin D1: a key endogenous inhibitor of neuroinflammation. Biofactors 2022; 48:1005-26. doi: 10.1002/biof.1891 [Crossref] [ Google Scholar]
- Dartt DA, Hodges RR, Serhan CN. Immunoresolvent resolvin D1 maintains the health of the ocular surface. Adv Exp Med Biol 2019; 1161:13-25. doi: 10.1007/978-3-030-21735-8_3 [Crossref] [ Google Scholar]
- Spinosa M, Su G, Salmon MD, Lu G, Cullen JM, Fashandi AZ. Resolvin D1 decreases abdominal aortic aneurysm formation by inhibiting NETosis in a mouse model. J Vasc Surg 2018; 68:93S-103S. doi: 10.1016/j.jvs.2018.05.253 [Crossref] [ Google Scholar]
- Xu J, Duan X, Hu F, Poorun D, Liu X, Wang X. Resolvin D1 attenuates imiquimod-induced mice psoriasiform dermatitis through MAPKs and NF-κB pathways. J Dermatol Sci 2018; 89:127-35. doi: 10.1016/j.jdermsci.2017.10.016 [Crossref] [ Google Scholar]
- Carion TW, Kracht D, Strand E, David E, McWhirter C, Ebrahim AS. VIP modulates the ALX/FPR2 receptor axis toward inflammation resolution in a mouse model of bacterial keratitis. Prostaglandins Other Lipid Mediat 2019; 140:18-25. doi: 10.1016/j.prostaglandins.2018.12.001 [Crossref] [ Google Scholar]
- Erdinest N, Ovadia H, Kormas R, Solomon A. Anti-inflammatory effects of resolvin-D1 on human corneal epithelial cells: in vitro study. J Inflamm (Lond) 2014; 11:6. doi: 10.1186/1476-9255-11-6 [Crossref] [ Google Scholar]
- Petrillo F, Trotta MC, Bucolo C, Hermenean A, Petrillo A, Maisto R. Resolvin D1 attenuates the inflammatory process in mouse model of LPS-induced keratitis. J Cell Mol Med 2020; 24:12298-307. doi: 10.1111/jcmm.15633 [Crossref] [ Google Scholar]
- Zhou J, Wu K, Ma Y, Zhu J, Zhou Y, Zhang Z. GTS-21 alleviates sepsis-induced atrial fibrillation susceptibility by modulating macrophage polarization and neuregulin-1 secretion. Int Immunopharmacol 2025; 154:114561. doi: 10.1016/j.intimp.2025.114561 [Crossref] [ Google Scholar]
- Deng Y, Li B, Zheng H, Liang L, Yang Y, Liu S. Multifunctional Prussian blue nanoparticles loading with Xuetongsu for efficient rheumatoid arthritis therapy through targeting inflammatory macrophages and osteoclasts. Asian J Pharm Sci 2025; 20:101037. doi: 10.1016/j.ajps.2025.101037 [Crossref] [ Google Scholar]
- Gong Z, Liu M, Liu H, Deng Z, Qin X, Nie J. Structural features and in vitro antitumor activity of a water-extracted polysaccharide from Ganoderma applanatum. New J Chem 2023; 47:13205-17. doi: 10.1039/d3nj01903a [Crossref] [ Google Scholar]
- Zhang Y, Zhu L, Li X, Ge C, Pei W, Zhang M. M2 macrophage exosome-derived lncRNA AK083884 protects mice from CVB3-induced viral myocarditis through regulating PKM2/HIF-1α axis mediated metabolic reprogramming of macrophages. Redox Biol 2024; 69:103016. doi: 10.1016/j.redox.2023.103016 [Crossref] [ Google Scholar]
- Wang X, Xie N, Zhang H, Zhou W, Lei J. Isoorientin ameliorates macrophage pyroptosis and atherogenesis by reducing KDM4A levels and promoting SKP1-Cullin1-F-box E3 Ligase-mediated NLRP3 ubiquitination. Inflammation 2025. doi: 10.1007/s10753-025-02289-2.
- Gao H, Wang L, Lyu Y, Jin H, Lin Z, Kang Y. The P2X7R/NLRP3 inflammasome axis suppresses enthesis regeneration through inflammatory and metabolic macrophage-stem cell cross-talk. Sci Adv 2025; 11:eadr4894. doi: 10.1126/sciadv.adr4894 [Crossref] [ Google Scholar]
- Ma W, Wei S, Peng W, Sun T, Huang J, Yu R. Antioxidant effect of Polygonatumsibiricum polysaccharides in D-galactose-induced heart aging mice. Biomed Res Int 2021; 2021:6688855. doi: 10.1155/2021/6688855 [Crossref] [ Google Scholar]
- Li WQ, Tan SL, Li XH, Sun TL, Li D, Du J. Calcitonin gene-related peptide inhibits the cardiac fibroblasts senescence in cardiac fibrosis via up-regulating klotho expression. Eur J Pharmacol 2019; 843:96-103. doi: 10.1016/j.ejphar.2018.10.023 [Crossref] [ Google Scholar]
- Qiao H, Ren H, Liu Q, Jiang Y, Wang Q, Zhang H, et al. Anti-inflammatory effects of Rehmanniaglutinosa polysaccharide on LPS-induced acute liver injury in mice and related underlying mechanisms. J Ethnopharmacol2025. 351: 120099. doi: 10.1016/j.jep.2025.120099.
- Van den Bossche J, Baardman J, Otto NA, van der Velden S, Neele AE, van den Berg SM. Mitochondrial dysfunction prevents repolarization of inflammatory macrophages. Cell Rep 2016; 17:684-96. doi: 10.1016/j.celrep.2016.09.008 [Crossref] [ Google Scholar]
- Davies J, Davies D. Origins and evolution of antibiotic resistance. Microbiol Mol Biol Rev 2010; 74:417-33. doi: 10.1128/mmbr.00016-10 [Crossref] [ Google Scholar]
- Wu J, Gao J, Yi L, Gao N, Wang L, Zhu J. Protective effects of resolvin D1 in Pseudomonas aeruginosa keratitis. Mol Immunol 2023; 158:35-42. doi: 10.1016/j.molimm.2023.04.007 [Crossref] [ Google Scholar]
- Luan S, Peng X, Lin J, Zhang Y, Zhan L, Yin J. Gallic acid ameliorates Aspergillus fumigatus keratitis through reducing fungal load and suppressing the inflammatory response. Invest Ophthalmol Vis Sci 2022; 63:12. doi: 10.1167/iovs.63.12.12 [Crossref] [ Google Scholar]
- Jin Y, Arita M, Zhang Q, Saban DR, Chauhan SK, Chiang N. Anti-angiogenesis effect of the novel anti-inflammatory and pro-resolving lipid mediators. Invest Ophthalmol Vis Sci 2009; 50:4743-52. doi: 10.1167/iovs.08-2462 [Crossref] [ Google Scholar]
- Qin Q, Hu K, He Z, Chen F, Zhang W, Liu Y. Resolvin D1 protects against Aspergillus fumigatus keratitis in diabetes by blocking the MAPK-NF-κB pathway. Exp Eye Res 2022; 216:108941. doi: 10.1016/j.exer.2022.108941 [Crossref] [ Google Scholar]
- Kanangat S, Meduri GU, Tolley EA, Patterson DR, Meduri CU, Pak C. Effects of cytokines and endotoxin on the intracellular growth of bacteria. Infect Immun 1999; 67:2834-40. doi: 10.1128/iai.67.6.2834-2840.1999 [Crossref] [ Google Scholar]
- Cejka C, Cejkova J. Oxidative stress to the cornea, changes in corneal optical properties, and advances in treatment of corneal oxidative injuries. Oxid Med Cell Longev 2015; 2015:591530. doi: 10.1155/2015/591530 [Crossref] [ Google Scholar]
- Hazlett LD, McClellan SA, Barrett RP, Huang X, Zhang Y, Wu M. IL-33 shifts macrophage polarization, promoting resistance against Pseudomonas aeruginosa keratitis. Invest Ophthalmol Vis Sci 2010; 51:1524-32. doi: 10.1167/iovs.09-3983 [Crossref] [ Google Scholar]
- Dhanushkodi NR, Srivastava R, Coulon PA, Prakash S, Roy S, Bagnol D. Healing of ocular herpetic disease following treatment with an engineered FGF-1 is associated with increased corneal anti-inflammatory M2 macrophages. Front Immunol 2021; 12:673763. doi: 10.3389/fimmu.2021.673763 [Crossref] [ Google Scholar]
- Zhu S, Chen Y, Lao J, Wu C, Zhan X, Wu Y. Signaling lymphocytic activation molecule family-7 alleviates corneal inflammation by promoting M2 polarization. J Infect Dis 2021; 223:854-65. doi: 10.1093/infdis/jiaa445 [Crossref] [ Google Scholar]
- Lee S, Kim S, Park S, Lee J, Yu HS. Effect of resolvin D1 on experimental bacterial keratitis to prevent corneal scar. Graefes Arch Clin Exp Ophthalmol 2022; 260:3293-302. doi: 10.1007/s00417-022-05686-7 [Crossref] [ Google Scholar]
- Kim SY, Lee JE. Resolvin D1 inhibits corneal inflammation in Staphylococcus aureus keratitis. Ocul Immunol Inflamm 2023; 31:927-34. doi: 10.1080/09273948.2022.2070504 [Crossref] [ Google Scholar]
- Schmid M, Gemperle C, Rimann N, Hersberger M. Resolvin D1 polarizes primary human macrophages toward a proresolution phenotype through GPR32. J Immunol 2016; 196:3429-37. doi: 10.4049/jimmunol.1501701 [Crossref] [ Google Scholar]
- Shan K, Feng N, Cui J, Wang S, Qu H, Fu G. Resolvin D1 and D2 inhibit tumour growth and inflammation via modulating macrophage polarization. J Cell Mol Med 2020; 24:8045-56. doi: 10.1111/jcmm.15436 [Crossref] [ Google Scholar]
- Hiram R, Xiong F, Naud P, Xiao J, Sirois M, Tanguay JF. The inflammation-resolution promoting molecule resolvin-D1 prevents atrial proarrhythmic remodelling in experimental right heart disease. Cardiovasc Res 2021; 117:1776-89. doi: 10.1093/cvr/cvaa186 [Crossref] [ Google Scholar]
- Xiang SY, Ye Y, Yang Q, Xu HR, Shen CX, Ma MQ. RvD1 accelerates the resolution of inflammation by promoting apoptosis of the recruited macrophages via the ALX/FasL-FasR/caspase-3 signaling pathway. Cell Death Discov 2021; 7:339. doi: 10.1038/s41420-021-00708-5 [Crossref] [ Google Scholar]
- Deng F, Yan J, Lu J, Luo M, Xia P, Liu S. M2 macrophage-derived exosomal miR-590-3p attenuates DSS-induced mucosal damage and promotes epithelial repair via the LATS1/YAP/β-catenin signalling axis. J Crohns Colitis 2021; 15:665-77. doi: 10.1093/ecco-jcc/jjaa214 [Crossref] [ Google Scholar]
- Lisovsky A, Zhang DK, Sefton MV. Effect of methacrylic acid beads on the sonic hedgehog signaling pathway and macrophage polarization in a subcutaneous injection mouse model. Biomaterials 2016; 98:203-14. doi: 10.1016/j.biomaterials.2016.04.033 [Crossref] [ Google Scholar]
- Jia YC, Qiu S, Xu J, Kang QL, Chai YM. Docosahexaenoic acid improves diabetic wound healing in a rat model by restoring impaired plasticity of macrophage progenitor cells. PlastReconstr Surg 2020; 145:942e-50e. doi: 10.1097/prs.0000000000006739 [Crossref] [ Google Scholar]
- Montanaro M, Meloni M, Anemona L, Giurato L, Scimeca M, Izzo V. Macrophage activation and M2 polarization in wound bed of diabetic patients treated by dermal/epidermal substitute Nevelia. Int J Low Extrem Wounds 2022; 21:377-83. doi: 10.1177/1534734620945559 [Crossref] [ Google Scholar]
- Nishida M, Fujinaka H, Matsusaka T, Price J, Kon V, Fogo AB. Absence of angiotensin II type 1 receptor in bone marrow-derived cells is detrimental in the evolution of renal fibrosis. J Clin Invest 2002; 110:1859-68. doi: 10.1172/jci15045 [Crossref] [ Google Scholar]
- Das A, Segar CE, Hughley BB, Bowers DT, Botchwey EA. The promotion of mandibular defect healing by the targeting of S1P receptors and the recruitment of alternatively activated macrophages. Biomaterials 2013; 34:9853-62. doi: 10.1016/j.biomaterials.2013.08.015 [Crossref] [ Google Scholar]
- Chen Z, Wu C, Gu W, Klein T, Crawford R, Xiao Y. Osteogenic differentiation of bone marrow MSCs by β-tricalcium phosphate stimulating macrophages via BMP2 signalling pathway. Biomaterials 2014; 35:1507-18. doi: 10.1016/j.biomaterials.2013.11.014 [Crossref] [ Google Scholar]
- Spiller KL, Nassiri S, Witherel CE, Anfang RR, Ng J, Nakazawa KR. Sequential delivery of immunomodulatory cytokines to facilitate the M1-to-M2 transition of macrophages and enhance vascularization of bone scaffolds. Biomaterials 2015; 37:194-207. doi: 10.1016/j.biomaterials.2014.10.017 [Crossref] [ Google Scholar]
- Loi F, Córdova LA, Zhang R, Pajarinen J, Lin TH, Goodman SB. The effects of immunomodulation by macrophage subsets on osteogenesis in vitro. Stem Cell Res Ther 2016; 7:15. doi: 10.1186/s13287-016-0276-5 [Crossref] [ Google Scholar]
- Córdova LA, Loi F, Lin TH, Gibon E, Pajarinen J, Nabeshima A. CCL2, CCL5, and IGF-1 participate in the immunomodulation of osteogenesis during M1/M2 transition in vitro. J Biomed Mater Res A 2017; 105:3069-76. doi: 10.1002/jbm.a.36166 [Crossref] [ Google Scholar]
- Li T, Peng M, Yang Z, Zhou X, Deng Y, Jiang C. 3D-printed IFN-γ-loading calcium silicate-β-tricalcium phosphate scaffold sequentially activates M1 and M2 polarization of macrophages to promote vascularization of tissue engineering bone. Acta Biomater 2018; 71:96-107. doi: 10.1016/j.actbio.2018.03.012 [Crossref] [ Google Scholar]
- Zhou T, He C, Lai P, Yang Z, Liu Y, Xu H. miR-204-containing exosomes ameliorate GVHD-associated dry eye disease. Sci Adv 2022; 8:eabj9617. doi: 10.1126/sciadv.abj9617 [Crossref] [ Google Scholar]
- Chandler HL, Tan T, Yang C, Gemensky-Metzler AJ, Wehrman RF, Jiang Q. MG53 promotes corneal wound healing and mitigates fibrotic remodeling in rodents. Commun Biol 2019; 2:71. doi: 10.1038/s42003-019-0316-7 [Crossref] [ Google Scholar]
- Kovács KJ. c-Fos as a transcription factor: a stressful (re)view from a functional map. Neurochem Int 1998; 33:287-97. doi: 10.1016/s0197-0186(98)00023-0 [Crossref] [ Google Scholar]
- Liu H, Lorenzini PA, Zhang F, Xu S, Wong MS, Zheng J. Alternative splicing analysis in human monocytes and macrophages reveals MBNL1 as major regulator. Nucleic Acids Res 2018; 46:6069-86. doi: 10.1093/nar/gky401 [Crossref] [ Google Scholar]
- Sacks D, Baxter B, Campbell BCV, Carpenter JS, Cognard C, Dippel D. Multisociety consensus quality improvement revised consensus statement for endovascular therapy of acute ischemic stroke: from the American Association of Neurological Surgeons (AANS), American Society of Neuroradiology (ASNR), Cardiovascular and Interventional Radiology Society of Europe (CIRSE), Canadian Interventional Radiology Association (CIRA), Congress of Neurological Surgeons (CNS), European Society of Minimally Invasive Neurological Therapy (ESMINT), European Society of Neuroradiology (ESNR), European Stroke Organization (ESO), Society for Cardiovascular Angiography and Interventions. J VascIntervRadiol 2018; 29:441-53. doi: 10.1016/j.jvir.2017.11.026 [Crossref] [ Google Scholar]
- Kim JY, Park HJ, Um SH, Sohn EH, Kim BO, Moon EY. Sulforaphane suppresses vascular adhesion molecule-1 expression in TNF-α-stimulated mouse vascular smooth muscle cells: involvement of the MAPK, NF-κB and AP-1 signaling pathways. VasculPharmacol 2012; 56:131-41. doi: 10.1016/j.vph.2011.11.007 [Crossref] [ Google Scholar]
- Hop HT, Arayan LT, Huy TXN, Reyes AW, Vu SH, Min W. The key role of c-Fos for immune regulation and bacterial dissemination in Brucella infected macrophage. Front Cell Infect Microbiol 2018; 8:287. doi: 10.3389/fcimb.2018.00287 [Crossref] [ Google Scholar]
- Lee SB, Shin JS, Han HS, Lee HH, Park JC, Lee KT. Kaempferol 7-O-β-D-glucoside isolated from the leaves of Cudraniatricuspidata inhibits LPS-induced expression of pro-inflammatory mediators through inactivation of NF-κB, AP-1, and JAK-STAT in RAW 2647 macrophages. Chem Biol Interact 2018; 284:101-11. doi: 10.1016/j.cbi.2018.02.022 [Crossref] [ Google Scholar]
- Wang A, Al-Kuhlani M, Johnston SC, Ojcius DM, Chou J, Dean D. Transcription factor complex AP-1 mediates inflammation initiated by Chlamydia pneumoniae infection. Cell Microbiol 2013; 15:779-94. doi: 10.1111/cmi.12071 [Crossref] [ Google Scholar]
- Chen QW, Wu GD, Yan ZY, Zhang WC, Zhang M. [Differential expression of immediately early gene c-fos in THP-1 macrophage subtype polarization]. Zhongguo Ying Yong Sheng Li Xue Za Zhi 2020; 36:534-8. doi: 10.12047/j.cjap.5865.2020.113 [Crossref] [ Google Scholar]