Bioimpacts. 16:32651.
doi: 10.34172/bi.32651
Review
Current advancements and future prospects in the biorefinery and hyper-production of astaxanthin from Haematococcus pluvialis
Jafar Aghajani Conceptualization, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing, 1 
Morteza Mirzaei Supervision, Writing – review & editing, 1
Ali Choopani Conceptualization, Data curation, Writing – review & editing, 1
Hosna Abbasi Data curation, Investigation, Writing – review & editing, 2
Fatemeh Shakeri Investigation, Visualization, Writing – review & editing, 1
Ali Mohammad Latifi Conceptualization, Formal analysis, Project administration, Validation, Writing – review & editing, 1, *
Author information:
1Applied Biotechnology Research Center, New Health Technologies Institute, Baqiyatallah University of Medical Sciences, Tehran, Iran
2Department of Microbiology, Science and Research Branch, Islamic Azad University, Tehran, Iran
Abstract
Astaxanthin (AXT) is a valuable xanthophyll carotenoid extensively utilized across the nutraceutical, pharmaceutical, cosmetic, and aquaculture industries. While this high-demand compound is produced both synthetically and biologically, natural AXT derived from the microalga Haematococcus lacustris (formerly called Haematococcus pluvialis) offers superior bioavailability and safety profiles, dominating the high-value market segment. However, the current industrial reality is hampered by low yields under standard cultivation, significantly restricting commercial viability. This comprehensive review systematically summarizes the latest advancements (2020-2025) in achieving cost-effective and sustainable hyper-production of natural AXT from H. pluvialis. We critically evaluate both established methodologies (e.g., two-phase cultivation, stress induction) and emerging strategies, including CRISPR-Cas9 editing, electrogenetics, and the application of nano-based stressors. Recent studies highlight significant yield enhancements, with reports of 20-35% increases via gene knock-ins and mild electric stimulation, achieving concentrations up to 200 mg L⁻¹. Crucially, this work provides an integrative comparison of advanced genetic engineering tools, nano-based induction protocols, and hybrid cultivation systems, distinguishing it from previous reviews focused predominantly on environmental factor optimization. The analysis offers actionable, evidence-based insights for accelerating the transition from lab-scale innovation to scalable industrial bioprocesses, thereby reducing production costs and enhancing the overall sustainability of natural AXT supply.
Graphical Abstract
Keywords: Haematococcus pluvialis, Astaxanthin, Photobioreactors, Metabolic engineering, CRISPR-cas9, Biotechnological strategies
Copyright and License Information
© 2026 The Author(s).
This work is published by BioImpacts as an open access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (
http://creativecommons.org/licenses/by-nc/4.0/). Non-commercial uses of the work are permitted, provided the original work is properly cited.
Funding Statement
The authors declare that no funds, grants, or other support were received during the preparation of this manuscript. No funding was received for conducting this study.
Introduction
Scientific Importance and Health Benefits of Astaxanthin
The use of chemicals combined with an excessive and erratic lifestyle can lead to the production of free radicals, which harm the human body. Antioxidants prevent or reduce the risk of diseases caused by excess free radicals. Carotenoids comprise over 750 naturally occurring pigments with strong antioxidant properties.1,2 Secondary carotenoids are produced in response to stress to maintain cellular redox balance. The potent antioxidant capacity of AXT has been highlighted in high-impact studies, demonstrating its role in mitigating oxidative stress across cellular systems.3,4
Astaxanthin (AXT) is one of the most potent antioxidants identified to date, particularly among carotenoids. AXT is a light-dependent carotenoid.5,6 Notably, xanthophylls may contain one or more oxygen-bearing functional groups, such as hydroxyl or ketone groups, whereas carotenes consist solely of hydrocarbons.7 AXT's molecular structure confers unique biochemical properties, including antioxidant and anti-inflammatory effects, owing to its hydroxyl and carbonyl functional groups, polar configuration, and esterification potential.8,9
AXT, also known as 3,3'-dihydroxy-β,β-carotene-4,4'-dione, is a naturally occurring xanthophyll, along with carotenes.10 The chemical structure of AXT is shown in Fig. 1. It contains two keto groups and two hydroxyl groups at positions 3,3' and 4,4' of the β-ionone rings, respectively. Its chemical formula is 4,4'-dione-β,β'-carotene-3,3'-dihydroxy-β.11 Additionally, the compound can readily cross cell membranes due to its lipophilic backbone flanked by hydrophilic groups at both ends. This process does not compromise the structural integrity or electron density of the bilayer membrane. Consequently, AXT exerts its antioxidant effects both intracellularly and extracellularly.12
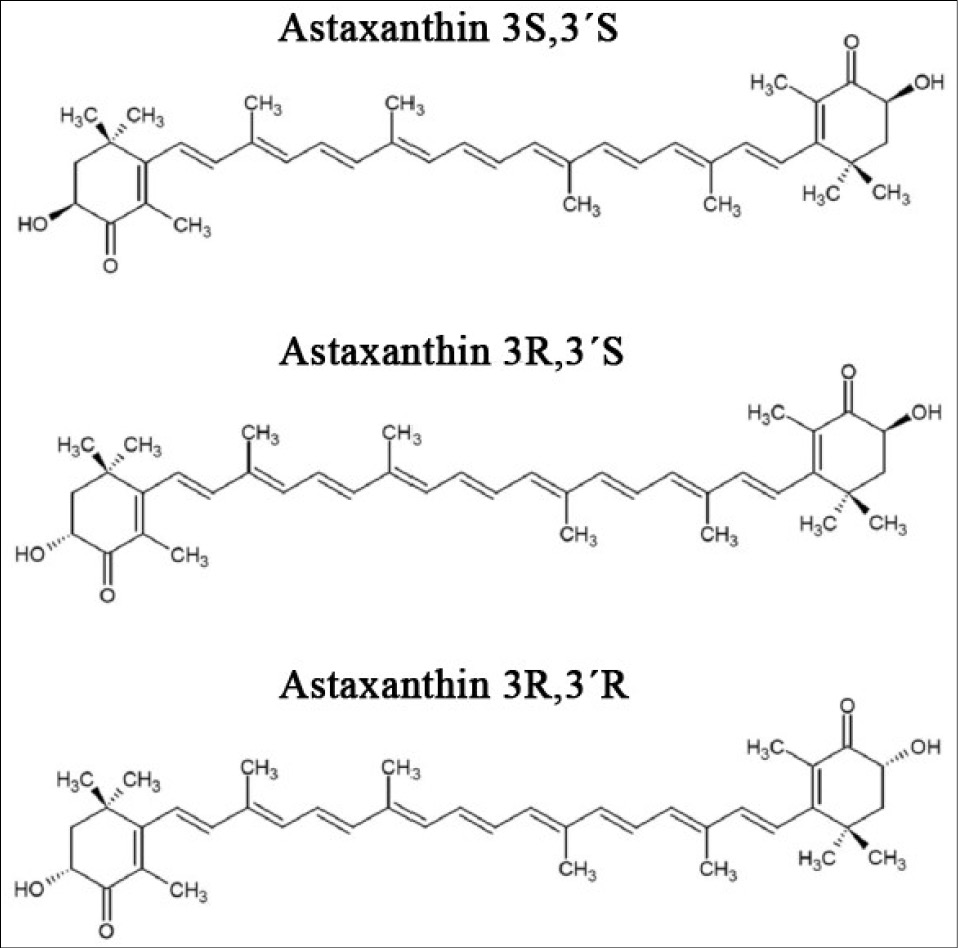
Fig. 1.
Structural formula of astaxanthin.
.
Structural formula of astaxanthin.
This carotenoid has multiple applications in human health and nutrition. It is particularly effective in treating various diseases by counteracting oxidative cellular damage, including cardiovascular, gastrointestinal, liver, and cancer-related conditions.6
The primary applications of AXT include food, medicine, nutraceuticals, and dietary supplements, primarily related to human nutrition and health.13,14 Its importance extends beyond these areas, as it is a significant commercial component in aquaculture, cosmetics, and health food supplements. This is mainly due to its unique reddish color and exceptional antioxidant capabilities.15,16 Moreover, AXT is a promising therapeutic agent with potential benefits for immune modulation, anti-aging, anti-cancer, and cardiovascular protection.9,17-20 Although AXT exhibits strong antioxidative activity (reported as up to 1000 times that of vitamin E, 200 times higher than tea polyphenols, 17 times higher than grape seed, and 10 times higher than other carotenoids like lutein, Canthaxanthin, β-carotene, and zeaxanthin in vitro)21-23 efficacy can vary in vivo due to bioavailability factors. Potential limitations include minimal side effects at high doses (e.g., skin pigmentation), as noted in safety reviews,24 but contradictory studies suggest variable anti-inflammatory effects in humans.17 Further studies are required to quantify its clinical advantages in humans. AXT and β-carotene from microalgae, including H. pluvialis, are suitable for human consumption owing to their natural origin and proven safety. Natural AXT from this alga has no known side effects and is designated GRAS (Generally Recognized as Safe) by the U.S. Food and Drug Administration (FDA). It is also registered by the Brazilian Health Surveillance Agency (ANVISA) and holds a prominent global market position.10,25 AXT from H. pluvialis has received regulatory approvals in several countries, including the United States, China, Japan, and various European nations, facilitating its use in dietary supplements and fortified foods.10
Sources and production methods
AXT can be produced via the more expensive biological process or the more economical chemical synthesis method. AXT is obtained from two main sources: natural and synthetic. Synthetic AXT, derived from petroleum derivatives, is primarily used in animal feed, and its application in human nutrition, cosmetics, and pharmaceuticals is not recommended due to potential health risks. In contrast, natural production aligns with biological principles and sustainable development. Therefore, direct consumption of synthetic AXT as a food supplement or drug for humans is not recommended.8,24,26
Biological sources of AXT encompass a diverse range of microorganisms. Microalgae such as Chlorella zofingiensis and Haematococcus pluvialis (H. pluvialis) are primary producers, whereas Chlorella sorokiniana, Tetraselmis sp., and Scenedesmus sp. serve as secondary sources.11,27 Yeasts including Phaffia rhodozyma, Xanthophyllomyces dendrorhous, and Yarrowia lipolytica also produce AXT.28 Additionally, certain bacteria, such as Agrobacterium spp., Brevibacterium spp., and Paracoccus spp., synthesize this compound.28-30 Marine organisms, including Aurantiochytrium, are also natural sources of AXT.31
The single-celled alga H. pluvialis, a member of the Chlorophyceae class and found in aquatic environments worldwide, is the premier natural source for commercial AXT production. It can accumulate up to 5% of its dry weight as AXT (primarily in esterified form), rendering it valuable for industrial applications.9,11,32,33 H. pluvialis, belonging to the phylum Chlorophyta, contains photosynthetic pigments such as chlorophyll a and b, along with various carotenoids. AXT constitutes approximately 90% of its total carotenoids, indicating a high pigment concentration in the dry biomass.34 This alga can be cultivated autotrophically (using light as an energy source) or heterotrophically (using organic carbon sources). Owing to its high AXT yield and superior biological properties, H. pluvialis is the most competitive natural source for industrial AXT production compared to other microbial producers.11
H. pluvialis renowned for its remarkable ability to produce AXT, a valuable carotenoid, particularly during the transition from motile vegetative cells to red dormant cysts. This transformation occurs under stress conditions, such as nutrient deprivation or high light intensity, resulting in rapid and substantial AXT accumulation as a defense against oxidative stress.11,35
H. pluvialis has a complex life cycle comprising two main phases (motile and nonmotile) and several stages, including microzooids, macrozooids, palmelloids, aplanospores, and hematocysts (Fig. 2).10 Under favorable conditions, flagellated cells (macrozooids) measuring 20–50 μm predominate. Under environmental stresses such as nutrient deficiency or high light intensity, these cells lose their flagella and transform into nonmotile forms (palmelloids), followed by red aplanospores or hematocysts.13,36,37 AXT accumulation begins in the intermediate stage, marked by a green-orange color change over 7–10 days (Fig. 2c), and peaks in the aplanospore stage. These structural and biochemical changes make H. pluvialis an ideal source for natural AXT production (Fig. 2).
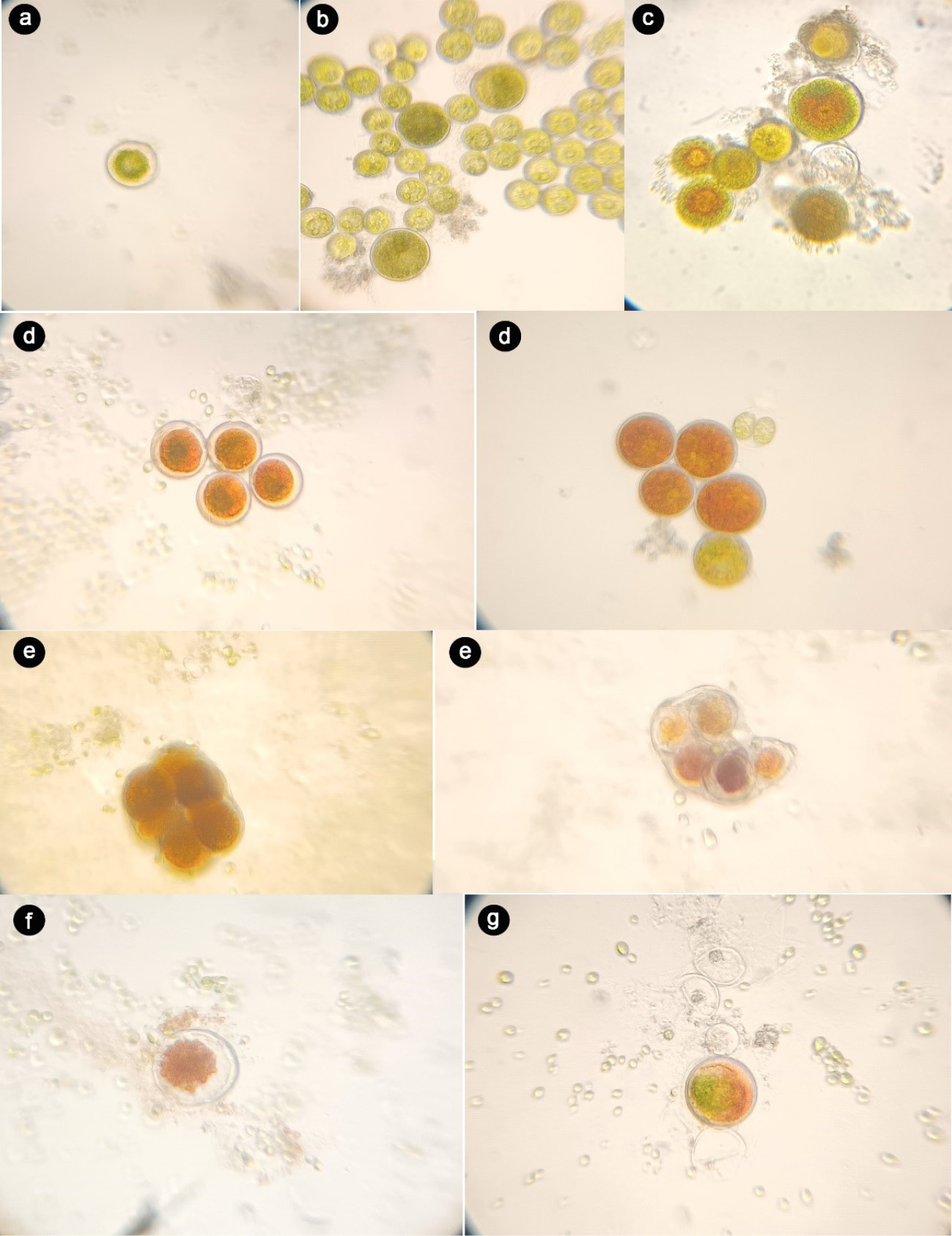
Fig. 2.
Cell cycle of H. pluvialis: (a) green vegetative motile cell (flagellated), (b) aplanospore, (c) astaxanthin-accumulating aplanospore, (d) red cyst cell, (e) sporangium, (f) astaxanthin rich zoospores released from the sporangium, (g) aplanospore derived from astaxanthin containing zoospore. Scale bar: 10 µm.
.
Cell cycle of H. pluvialis: (a) green vegetative motile cell (flagellated), (b) aplanospore, (c) astaxanthin-accumulating aplanospore, (d) red cyst cell, (e) sporangium, (f) astaxanthin rich zoospores released from the sporangium, (g) aplanospore derived from astaxanthin containing zoospore. Scale bar: 10 µm.
In the vegetative phase, H. pluvialis cells contain 26–60% protein, 25–39% carbohydrate, and 8–23% lipid on a dry weight basis. Upon entering the cyst phase, the biochemical composition changes significantly: protein decreases to 12–14%, carbohydrate increases to 43–74%, and lipid to 35–40%. Carotenoid content also increases to 2–5% of dry weight, which AXT comprising 80–99%. This biochemical shift not only aids algal survival under adverse conditions but also enhances its commercial value as an AXT source.38
Most companies employ a two-phase approach for cultivating H. pluvialis to produce AXT.39-41 This strategy is based on the alga's life cycle and cellular properties, comprising at least two distinct phases: the green stage (growth phase), where cells multiply under favorable conditions to increase biomass; and the red stage (non-motile phase), where cells are exposed to stress factors to induce AXT synthesis.36 In the green stage, cells reproduce and accumulate biomass but do not produce AXT. In contrast, during the red stage, they lose reproductive and motile capabilities but can accumulate AXT up to 5% of dry biomass weight.39-41 In the initial phase, culture conditions are optimized to maintain vegetative cells for robust division. Once target cell density is reached, physical or chemical adjustments activate AXT metabolic pathways (Fig. 3). This workflow (Fig. 3) integrates recent advancements in electrotechnologies for cell disruption, as demonstrated by Gherabli and colleagues.42
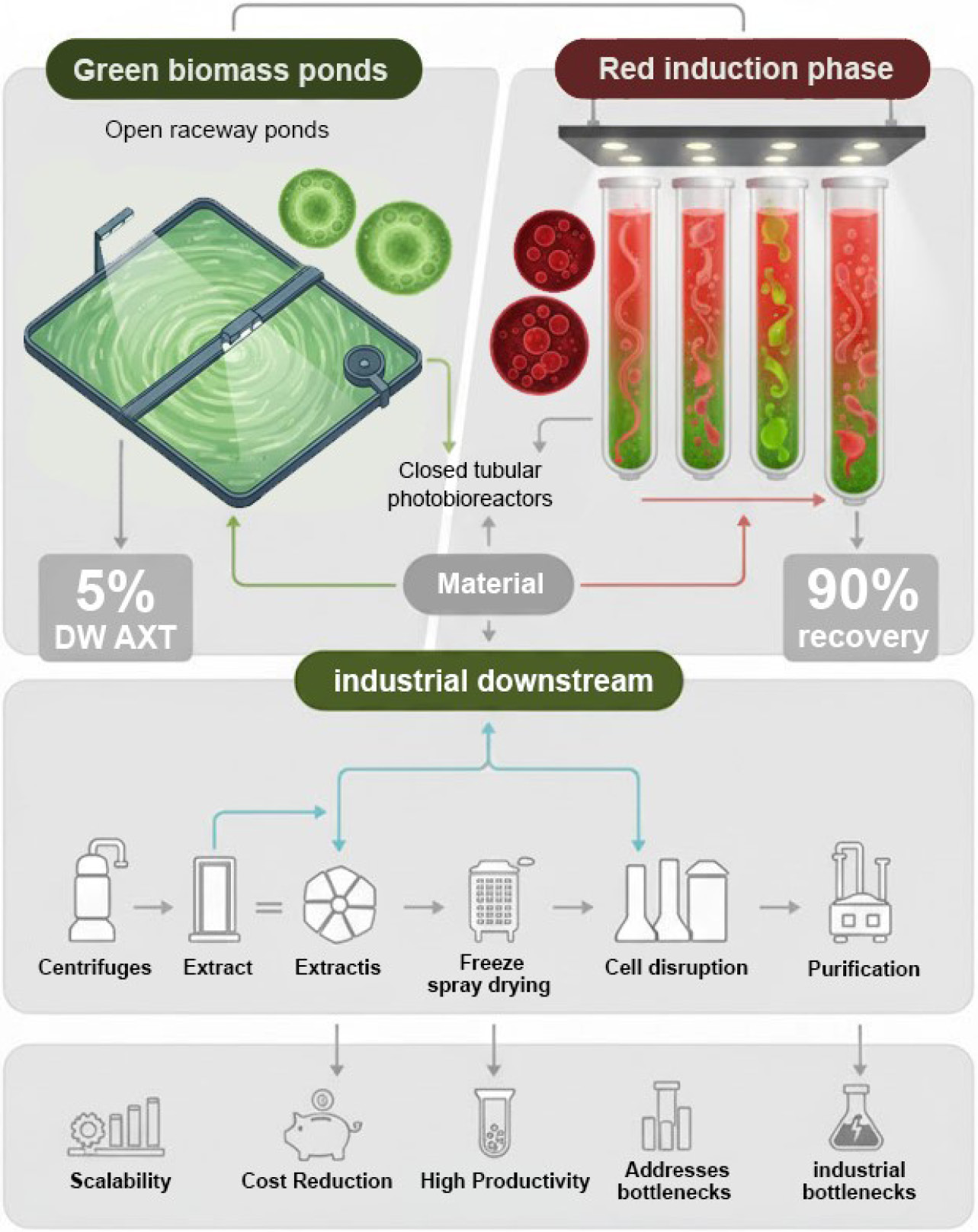
Fig. 3.
Industrial downstream processing workflow for astaxanthin extraction from H. pluvialis biomass obtained via a two-phase approach for cultivating. The schematic illustrates a sequential process commencing with biomass harvesting through sedimentation or centrifugation, followed by freeze or spray drying, cell wall disruption via mechanical, enzymatic, or electrical methods (e.g., pulsed electric fields), solvent or supercritical fluid extraction, and purification by chromatography to yield the final product. Adapted from concepts in Refs.40,42,43
.
Industrial downstream processing workflow for astaxanthin extraction from H. pluvialis biomass obtained via a two-phase approach for cultivating. The schematic illustrates a sequential process commencing with biomass harvesting through sedimentation or centrifugation, followed by freeze or spray drying, cell wall disruption via mechanical, enzymatic, or electrical methods (e.g., pulsed electric fields), solvent or supercritical fluid extraction, and purification by chromatography to yield the final product. Adapted from concepts in Refs.40,42,43
Thus, balancing the green and red stages is essential for maximizing yields. In this two-step process, H. pluvialis cells are first cultivated in photobioreactors under standard conditions to promote proliferation and biomass accumulation. Subsequently, they are transferred to larger photobioreactors or raceway ponds under stressful, nutrient-limited conditions to enhance AXT accumulation and encystment.8,9,30 AXT extraction from microalgal cultivation involves biomass harvesting, drying, cell disruption, and isolation. Commercially, centrifugation is the most common harvesting technique for H. pluvialis, often preceded by passive sedimentation due to the cells’ high density (Fig. 4).40,42
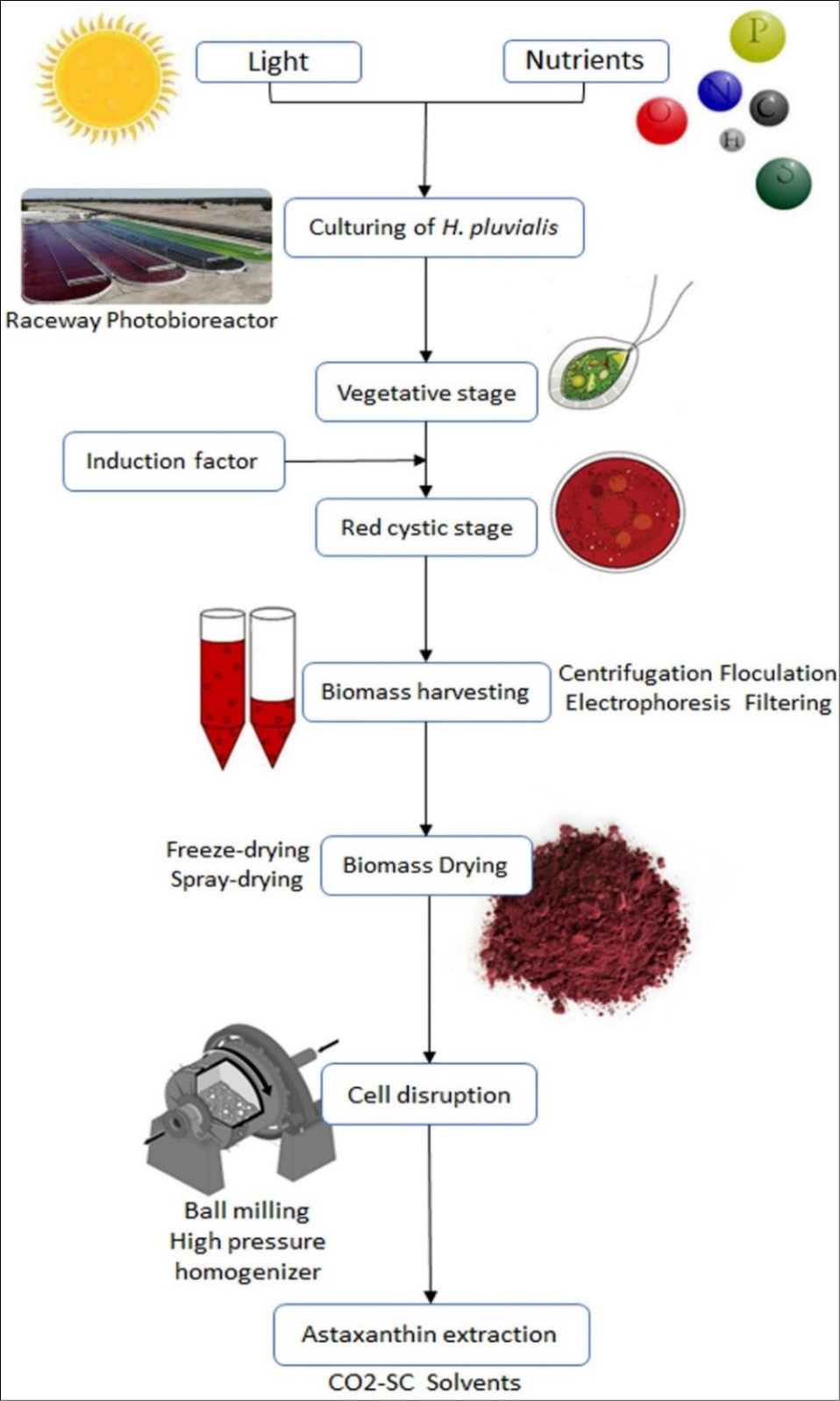
Fig. 4.
Integrated bioprocess pipeline for the production and extraction of astaxanthin from the microalga Haematococcus pluvialis. The process begins with large-scale cultivation (Raceway Photobioreactor) of the vegetative stage, followed by the induction of the astaxanthin-rich red cystic stage using stress factors. The resulting biomass is harvested, dried, and subsequently subjected to cell disruption and final astaxanthin extraction using methods like CO2 Supercritical fluid (SC) to obtain the high-value carotenoid.
.
Integrated bioprocess pipeline for the production and extraction of astaxanthin from the microalga Haematococcus pluvialis. The process begins with large-scale cultivation (Raceway Photobioreactor) of the vegetative stage, followed by the induction of the astaxanthin-rich red cystic stage using stress factors. The resulting biomass is harvested, dried, and subsequently subjected to cell disruption and final astaxanthin extraction using methods like CO2 Supercritical fluid (SC) to obtain the high-value carotenoid.
Enhanced AXT production in H. pluvialis can be achieved through stress induction, including high light intensity, salinity and pH changes, electrical stimulation, nuclear irradiation, nutrient deprivation, carbon-to-nitrogen ratio adjustments, organic carbon addition, and chemical agents. These methods optimize the AXT yields for industrial applications by mimicking natural stresses (Table 1).36,44-49
Table 1.
Methods for inducing astaxanthin production, including stress types and mechanisms
|
Induction Method
|
Explanation
|
References
|
| Intense light |
Stimulating astaxanthin production with high-intensity light |
44,50,51
|
| Salinity changes |
Increasing or decreasing salinity to create stress |
44,52-55
|
| PH adjustment |
Changing the pH of the culture medium as a stressor |
44,56
|
| Electrical stimulation |
Using electrical currents to stimulate biosynthesis |
15,16,45,47,57
|
| Nuclear irradiation |
Creating beneficial mutations with nuclear radiation |
46,48
|
| Nutrient deficiency |
Limiting nitrogen or phosphorus to increase production |
10,47,58
|
| Adjusting carbon /nitrogen ratio |
Optimizing the ratio to improve production efficiency |
47
|
| Organic carbon sources |
Adding glucose or other carbon sources as an energy source |
36
|
| Chemical agents |
Using substances such as salicylic acid to stimulate production |
59
|
Industrial details and market overview
Currently, most industrial-scale AXT production relies on chemical methods using petrochemicals and their derivatives.60 Due to lower production costs and prices, synthetic AXT meets aquaculture market needs and supplies approximately 95% of the total AXT market. As opposed to this, natural AXT, although produced in smaller quantities, commands significantly higher prices due to its superior bioavailability and safety profile.32 Importantly, chemical synthesis produces molecules with various configurations, lacking many biological characteristics of natural AXT. Owing to its petrochemical origin and stereoisomers that may compromise molecular stability, synthetic AXT poses potential toxicity and carcinogenicity risks. Consequently, its primary application is in salmon feed for muscle pigmentation.60,61
Synthetic AXT, derived from petroleum derivatives, is primarily used in animal feed, and its application in human nutrition, cosmetics, and pharmaceuticals is not recommended due to potential health risks. In contrast, natural production aligns with biological principles and sustainable development.
Currently, synthetic AXT dominates the global market. However, reports of adverse effects from synthetic AXT consumption in humans have substantially increased demand for natural AXT.25 Projections indicate significant growth in the natural AXT market in the coming years. Natural AXT exhibits higher esterification levels (up to 95%) than its synthetic counterpart, enhancing energy production and protecting tissues from oxidative damage under stress. In contrast, synthetic AXT is poorly absorbed due to its crystalline structure, whereas natural AXT, being non-crystalline and lipid-soluble, shows superior absorption.62 Consequently, the retail price of natural nutraceutical-grade AXT exceeds US $100,000 per kilogram. In the market, natural AXT extracted from sources such as microalgae and yeast competes with the synthetic form, which dominates industrial production. However, synthetic AXT has lower market value, limited biological functions, usage restrictions, and greater environmental impacts. The global market for natural AXT is expected to reach US $1 billion by 2025, driven by growing demand in sectors such as aquaculture, food, cosmetics, and pharmaceuticals.8,9,63 This surge has attracted new producers and garnered attention from various sectors. AXT prices typically ranges from $2,000 to $15,000 per kilogram, depending on purity, sales location, and extraction source (natural or synthetic). Natural AXT from sources such as algae is generally more expensive than synthetic forms derived from petroleum derivatives.64
The primary applications of AXT include food, medicine, nutraceuticals, and dietary supplements, primarily related to human nutrition and health.13,14 Its importance extends beyond these areas, as it is a significant commercial component in aquaculture, cosmetics, and health food supplements. This is mainly due to its unique reddish color and exceptional antioxidant capabilities.15,16 It is also commonly applied as a colorant in the food and aquaculture industries.32
The H. pluvialis market is currently valued at approximately US $240 million, with raceway pond cultivation accounting for US $200 million and photobioreactors for US $40 million.65 This disparity arises from differences in the molecular structure of yeast-derived AXT, which has reduced biological efficacy, stricter usage restrictions, and lower suitability for human consumption. Table 2 provides a concise overview of the current and projected market outlook, highlighting key economic drivers and constraints.
Table 2.
Global Market Overview and Economic Parameters for H. pluvialis-Derived AXT (2024-2032)
|
Economic parameter
|
Details/Value
|
| Market value (2024) |
US $127 million |
| Projected value (2032) |
US $266 million |
| Projected CAGR (2025-2032) |
11.4% |
| Largest application segment (2024) |
Aquaculture ( > 35% of demand) |
| Fastest growing segment |
Cosmetics (Driven by UV-protection/anti-aging) |
| Production cost challenge |
Natural astaxanthin COGS must compete with synthetic astaxanthin (approx. 60-70% lower in price) |
CAGR: Compound Annual Growth Rate, COGS: Cost of Goods Sold.
Gaps and research objectives
Although production costs remain high, they can be reduced through more efficient cultivation and processing techniques for H. pluvialis biomass, combined with a biorefinery approach that generates valuable products and co-products.28,66 However, microalgal AXT production faces several challenges, including longer cultivation times (10–15 days) compared to yeasts and bacteria (7–10 days), slower growth rates, limited biomass yields, and lower carotenoid levels under standard conditions. Additionally, microalgae are susceptible to degradation or cellular collapse under stress, exhibit moderate AXT accumulation, and incur high cultivation costs. These factors continue to hinder economically viable industrial production of H. pluvialis.28,66
This review examines existing methodologies for producing natural AXT from H. pluvialis and proposes research directions and innovations to achieve technological advancements, ultimately reducing industrial production costs. It also offers practical recommendations for refining these processes. Conventional and advanced cultivation techniques for H. pluvialis are outlined, focusing on strategies for the green and red growth phases. These approaches encompass various conditions and methods to enhance biomass and AXT yields in H. pluvialis. Consequently, this review provides new insights into achieving high biomass concentrations and optimal AXT accumulation for industrial applications.
Various approaches to enhancing biomass and the accumulation of Astaxanthin
Numerous studies have investigated strategies to enhance biomass and AXT accumulation in H. pluvialis.40,46,62,67-69 This section examines key factors influencing AXT production in both stages: the green stage, focuses on cultivating green H. pluvialis cells to increase biomass, and the red stage, where stress is applied to induce AXT synthesis (Table 3). Table 3 comprehensively details physical, chemical, and advanced biotechnological approaches used to manipulate the microalgal metabolism, primarily by inducing stress conditions to shift the cell from the green vegetative stage to the AXT-rich red cystic stage, alongside genetic and electrogenetic methods for improved yield and productivity.
Table 3.
Key strategies employed for bioprocess optimization to enhance biomass and astaxanthin accumulation in H. pluvialis
|
Strategy category
|
Specific method
|
Mechanism of action
|
Impact on astaxanthin yield
|
Key findings/examples
|
Advantages
|
Challenges/Considerations
|
Source
|
| Environmental Factors (Stress Induction) |
Light Intensity |
Stimulates photosynthesis and Reactive Oxygen Species (ROS) production, shifting metabolism. |
Positive (High light is critical for accumulation). |
Low light is optimal for growth; high light induces accumulation. Constant light led to a 60% increase in astaxanthin. |
Precise control of cellular signaling and productivity increase. |
Requires complex photobioreactor design and precise control. |
65,70-72
|
| Different Light Colors |
Stimulates specific metabolic pathways via tailored wavelengths. |
Positive (e.g., Blue LED, Blue + UV). |
Blue LED enhances astaxanthin synthesis; Blue + UV led to a 4 × increase in chlorophyll and 23μg/cell carotenoid. |
Energy efficiency and precise physiological control. |
Requires advanced LED systems and a complex understanding of cellular response. |
5,73,74
|
| Temperature |
Directly affects photosynthetic machinery and cell division rate. |
Positive (Optimal range: 25−28∘C; specific 27∘C with N depletion). |
Maximum division at 20 °C. Synergy with N depletion at 27 °C resulted in a 37% increase in astaxanthin. |
Optimizes growth and accumulation; synergy with other factors (N depletion). |
Narrow optimal window; challenging to control at large-scale; risk of cell lysis at > 35-40 °C. |
70,75
|
| Salinity Changes |
Slows cell division and stimulates astaxanthin biosynthesis genes. |
Positive (e.g., 0.25−0.5%NaCl). |
NaCl with sodium acetate increases AXT accumulation and biosynthesis gene activity. |
Cost-effective; dual-purpose (stress induction and growth control); reduces contamination. |
High levels can cause cell bleaching and damage. |
52,76,77
|
| Nutrient Depletion (N and P) |
Shifts metabolic priority toward energy storage and
secondary synthesis (carotenoids). |
Potently Positive (especially N depletion). |
N depletion is more effective than high light. Limiting all nutrients except CO2 is also efficient. |
Most potent and cost-effective inducer for AXT synthesis. |
Requires precise nutrient management systems; negatively impacts overall biomass growth. |
58,72
|
| pH Adjustment |
Affects cell proliferation and metabolism. |
Not specified/Variable. |
H. pluvialis grows in a wide pH range (4.0−9.0). |
Reduces need for precise control and lowers operational costs. |
Direct effect as a sole induction factor for high AXT accumulation is minor; optimization is typically limited to the green stage. |
65,78
|
| Advanced Biotechnological Approaches |
Nanoparticles (NPs) |
Creates stress conditions via active radicals to stimulate AXT biosynthesis. |
Positive (at low concentrations). |
ZnO NPs and Mg NPs enhance AXT synthesis; Ag NPs promote peak carotenoid production. C-paints led to a 1.8 × AXT increase. |
Targeted stress induction; potential for increased productivity. |
Toxicity at high concentrations; reduced light penetration; requires precise dosing. |
79,80
|
| Strain Improvement (Mutagenesis) |
Alters genetic structure (random or induced) for desirable traits (e.g., enhanced growth or AXT production). |
Positive. |
Induced mutagenesis (UV, EMS, DPA) yielded a 1.7 × AXT increase and a 30% productivity boost in raceway ponds. |
Avoids GMO classification; significant improvements in yield and biomass. |
Requires efficient screening; some mutagens are toxic. |
71,81,82
|
| Genetic Modification (CRISPR-Cas9) |
Targeted genome editing to manipulate specific biosynthesis pathways (e.g., carotenoid pathway). |
Positive. |
In H. pluvialis, Cas9-Lag resulted in a 26.7% AXT and 29.0% zeaxanthin increase. |
High precision; allows specific pathway manipulation; significant yield increase. |
Regulatory challenges (GMO status); high technical complexity. |
4,83-85
|
| Electric Stimulation (Electrogenetics) |
Induces controlled ROS (via electric current) to stimulate AXT biosynthesis. |
Positive (e.g., 10 mA current for 4hrs). |
10mA for 4hrs resulted in a 21.8%−34.9% AXT increase; 30mA for 2days increased AXT by 36.9%. |
Sustainable and chemical-free; precise and cost-effective method. |
Narrow "therapeutic window"; risk of cell damage at high currents; requires precise monitoring. |
16,42,47,57
|
Various cultivation media
Researchers have evaluated various conventional media, including BBM, Z8, BAR, FAB, BG-11, KM1, MM1, and MM2, for H. pluvialis cultivation. Among these, BBM provides the optimal environment for green cell growth.86
Effect of Light and Illumination Intensity
Light intensity and color are fundamental to AXT production.62,66 As the primary energy source for phototrophic cultivation, light is essential for cell development and AXT accumulation. Research shows a positive correlation between biomass, AXT productivity, and both light intensity and duration.50,51 Specifically, extended photoperiods and high intensities significantly enhance AXT accumulation, particularly in outdoor settings.50,51
High light intensity induces reactive oxygen species (ROS) formation, which in turn activates carotenoid biosynthesis as a protective mechanism, thereby enhancing AXT accumulation. However, excessively high illumination can damage photosystems, reducing biomass growth. Moderate light levels (100–150 µmol photons m-2 s-1) provide a balance between sustained growth and pigment induction, while transient high-light exposure is optimal for short-term AXT stimulation.51
Moderate light conditions are key for effective AXT induction. To optimize yields, low light intensities are applied initially in the first phase to promote microalgal proliferation until optimal cell density is reached, followed by high light intensity as stress in the second phase to induce AXT accumulation.70,87
In a detailed study, Torzillo et al88 examined the effects of varying light intensities on green H. pluvialis cell cultivation, analyzing biomass growth, chlorophyll dynamics, photochemical parameters, oxygen evolution, and respiration under 50–600 μmol m-2 s-1. Notably, no saturation point for vegetative growth was observed below 200 μmol m-2 s-1.88 Furthermore, Azizi et al34 showed that in a 5-L stirred-tank photobioreactor with constant light during the phototrophic stage, biomass and AXT concentrations increased by 50% and 60%, respectively, compared to BG-11 cultures under 100 μmol m-2 s-1.34 Additionally, Lv et al89 reported significant increases in metabolites essential for AXT biosynthesis with escalating light intensity.89
Different light colors
Selecting specific light wavelengths is one of the most effective tools to modulate microalgal metabolism and direct cellular energy flow toward the biosynthesis of valuable secondary metabolites such as AXT.74,90,91
Monochromatic LED light influences microalgal physiology, making it pivotal for optimizing cultivation.74,91 Studies indicate that blue LED light (380–470 nm) promotes AXT production with minimal impact on biomass. Exposure to blue LED light substantially increases AXT in dry biomass. Based on this, a 2004 method proposed red LED light for the green phase and blue for the red, AXT-rich phase, which was tested in 2016.73
Blue wavelengths induce the highest AXT accumulation, exceeding white and red light effects, due to reduced absorption by photosystem II pigment-protein complexes, minimizing photodamage and photoinhibition.73
In a 2015 study, Suyono et al5 induced oxidative stress with ultraviolet light to enhance H. pluvialis carotenoid content and examined blue, red, and white light effects on biomass.5 Their results showed that ultraviolet and visible light contribute to cell growth and carotenogenesis. Red-ultraviolet pairing yielded the highest biomass and a threefold chlorophyll increase.5 Blue-ultraviolet combination produced a fourfold per-cell chlorophyll increase, the highest total carotenoid (5 mg/L), and per-cell carotenoid (23 μg/cell).5
Effect of temperature
A recent study optimizing H. pluvialis cultivation temperatures for industrial applications showed that temperature profoundly affects the photosynthetic machinery of H. pluvialis cells.75 It also revealed that temperature influences both cell division rates and cell size, highlighting the cultures' sensitivity to temperature fluctuations.75 Notably, the examined strain exhibited peak division rates at 20 °C, but fastest biomass accumulation at up to 30.5 °C. Multiple studies confirm that the upper limit for effective carotenogenesis in H. pluvialis is 30 °C, with temperatures exceeding 35–40 °C causing cell lysis.10,39,51,70,92
Giannelli et al75 reported a 37% increase in AXT accumulation at 27 °C compared to 20 °C (156 mg L⁻¹ vs. 115 mg L⁻¹) under nitrogen deprivation, with combined high temperature and nitrogen stress enhancing yields.75 Additionally, Hong et al93 found that 25–28 °C increased AXT production relative to 23 °C. They noted that 30–36 °C heat stress suppressed photosynthesis, with intracellular AXT levels negatively correlated to photosynthetic performance.93
Effect of pH
The pH of the culture medium is a critical physicochemical parameter influencing both vegetative growth (biomass production) and AXT induction (encystment) in H. pluvialis. The optimal pH range must be carefully maintained to maximize overall productivity, as extreme deviations can cause irreversible cellular damage and loss of culture viability. During the initial green growth phase, H. pluvialis exhibits maximal cell proliferation within a neutral to slightly alkaline range, typically between pH 7.0 and 8.0.94-96 Within this range, the microalga effectively utilizes dissolved inorganic carbon sources, primarily bicarbonate (
) and dissolved CO2. Deviations below pH 6.0 or above pH 9.0 significantly impede growth and may lead to cell lysis.96 For instance, published quantitative data show that while cell growth remains relatively stable from pH 5.0 to 9.0, the maximum cell density is generally achieved around a neutral pH.94 However, recent studies demonstrate that this microalga can thrive across a broader pH range (4.0–9.0), showing greater adaptability than previously assumed.56
In the red induction phase, where AXT accumulation is triggered, the cellular response to pH becomes more complex, often interacting with other stressors. Some studies suggest that a slightly acidic pH may enhance carotenogenesis. Research exploring the influence of pH during the induction stage found that an environment of pH 6.5 resulted in a superior AXT accumulation effect compared to other tested pH environments.97 This suggests that a minor shift toward acidity during the stress phase might optimize the cellular environment for enzymatic activity related to carotenoid biosynthesis, possibly by modulating the activity of key enzymes like pH-sensitive beta-carotene hydroxylase. Therefore, efficient large-scale cultivation protocols mandate precise pH control, often relying on automated pH controllers and CO2 supplementation to maintain the optimal range for each distinct production stage.
Salinity
Salinity has a complex effect on lipid productivity in microalgae such as H. pluvialis.52 Evidence shows that combining sodium chloride (NaCl) with 2.2 mM sodium acetate markedly increases AXT accumulation.52
NaCl is often used to retard cell division while promoting AXT production. At moderate concentrations (0.25%-0.5% w/v), NaCl effectively enhances AXT synthesis, but higher levels may cause cell bleaching under phototrophic and mixotrophic conditions.52,53,55 Salinity stress also upregulates genes involved in AXT biosynthesis.55 Incorporating NaCl (a cost-effective and widely available substance) into the culture medium can enhance biomass, lipid, and carotenoid yields while reducing AXT production costs.
Gao et al55 reported strain-specific salinity tolerance in H. pluvialis; for example, three Australian isolates grew at NaCl concentrations up to 0.17 M (∼1%).54 Separately, Tam et al54 investigated NaCl effects on H. pluvialis, finding that high salinity suppressed growth but increased per-cell carotenoid content, highlighting the trade-off between growth and AXT production under salinity stress.54
Nitrogen depletion strategy
Research shows that nutrient scarcity, especially nitrogen stress, significantly enhances intracellular carotenoid accumulation, including AXT, in microalgal cultures. Nitrogen deficiency has a greater impact on cost-effective AXT production than light intensity.10 Depleting both nitrogen and phosphorus rapidly promotes AXT accumulation and cellular encystment, with nitrogen depletion being more effective than phosphorus depletion.58 Moreover, restricting all nutrients except CO₂ may be the most efficient and economical approach for autotrophic AXT production, given the negligible difference in accumulation rates between nitrogen-specific and complete nutrient deprivation.58
Nitrogen deficiency limits protein synthesis, reducing photosynthetic efficiency and redirecting carbon flux toward secondary metabolites such as carotenoids. The accumulation of ROS during nutrient stress acts as a signaling trigger for transcriptional upregulation of key carotenogenic genes, including bkt and crtZ, leading to enhanced AXT biosynthesis.58 Nitrogen is more critical than phosphorus due to its role in amino acid biosynthesis, halting cell division faster than phosphorus limitation.10
Nanoparticles
In recent years, researchers have investigated various nanoparticles (NPs) to increase AXT yields in microalgae by inducing radical-driven stress.80,98,99 For example, zinc oxide NPs amplify AXT levels by stimulating cell proliferation and content.100
Nanoparticles (e.g., ZnO, TiO₂) generate free radicals via surface catalysis (interaction with cellular membranes and chloroplasts), elevating ROS levels that activate mitogen-activated protein kinase (MAPK) and nuclear factor erythroid 2-related factor 2 (NRF2) signaling pathways, upregulating genes like pds and lcy -b for AXT biosynthesis.100 This stress mimics environmental cues, increasing yields by 1.8-fold at 1 mg/mL.98
Further studies examined silver NPs (SNPs) at 10, 100, and 200 ppm for AXT biosynthesis; initially slowing growth, they increased biomass after acclimation, with peak carotenoid production at 10 ppm.99 Similarly, H. pluvialis exposure to biocompatible liquid fluorescent carbon nanodots (C-paints) at 1 mg/mL yielded a 1.8-fold AXT increase compared to controls.98 Likewise, the application of magnesium nanoparticles has been found to enhance AXT synthesis in H. pluvialis.80 Rastar et al100 reported that zinc NPs at 2.49 and 4.41 mg/L in BBM medium increased biomass, AXT, and chlorophyll levels, aligning with standard formulations.100 However, high metal NP concentrations can cause toxicity, reducing cell density due to impaired light penetration and photosynthesis.100
Strain improvement by mutagenesis
Mutagenesis provides a powerful approach to modify the genetic structure and properties of microalgal strains, enhancing their industrial potential similar to methods used for traditional industrial microorganisms. Using genetic tools like gene expression and editing, endogenous and exogenous genes can be tailored to specific goals. Random mutagenesis is a key technique for microalgal strain improvement.59,101,102
Critical genes in AXT biosynthesis (such as lycopene β-cyclase, phytoene desaturase, β-carotene oxygenase, β-ring hydroxylase, β-carotene ketolase, and ζ-carotene desaturase) are central to this process. Induced mutagenesis strengthens inheritable microalgal traits, and diverse editing strategies enable targeted mutagenesis, fostering novel genotypes.59,101,102
Mutagenesis occurs naturally but can be induced in laboratories using physical or chemical agents, even without comprehensive genome knowledge. Agent-driven lab mutagenesis mimics natural pathways, producing desirable mutants not classified as Genetically Modified Organisms (GMOs), thus facing fewer regulatory hurdles.103
Chemical mutagens like ethyl methanesulfonate are effective for random mutagenesis to amplify AXT production genetically. Similarly, N-methyl-N'-nitro-N-nitrosoguanidine (MNNG) is a potent mutagen for H. pluvialis, generating cell wall-deficient mutants with higher AXT yields.104 Gomez et al demonstrated chemical mutagenesis combined with herbicide selection to improve strain traits for industrial use.105 Starting with a wild-type Chilean strain, they generated mutants; one grown in 25,000-L outdoor raceway ponds achieved 30% higher productivity than the wild type.105
Wang et al71 used a three-stage strategy with UV irradiation, ethyl methanesulfonate (EMS), and diphenylamine (DPA), increasing cell dry weight, extending logarithmic growth, and identifying high-AXT mutants. The DPA12-2 mutant showed a 1.7-fold yield increase over the wild type.71 Cheng et al applied48 Co-γ irradiation under high CO2, raising AXT to 46.0 mg/L and lipid content by 45.9%.59
Early techniques used UV radiation and mutagens like EMS and diethyl sulfate (DES). Herbicides such as DPA and nicotine served as selective agents by inhibiting carotenoid synthesis, yielding mutants with elevated AXT and lycopene cyclase activity compared to wild types under lab conditions.106,107
Liu et al used Fourier transform infrared (FT-IR) and Raman microspectroscopy for non-invasive AXT quantification in cells.108 They identified a strong correlation between the FT-IR band ratio I(1740)/I(1156) and single-cell AXT content, enabling screening for hyper-producing mutants.108 In 2018, Hong et al exposed UV-mutated H. pluvialis to sodium azide (an antioxidant inducer), producing improved mutants.102 Among various mutants screened, the most productive strain (M13) exhibited an 88.5% increase in AXT content relative to the wild-type culture, indicating a substantial enhancement in carotenoid synthesis efficiency rather than the proportion of mutants generated.102
Various successful strain improvement methodologies utilizing physical and chemical mutagens, along with specific selection techniques, are comparatively detailed, highlighting their quantitative outcomes in Table 4.
Table 4.
Comparative Summary of Strain Improvement Methods for Enhanced Astaxanthin Production in H. pluvialis
|
Mutagenesis method/agent
|
Strain selection mechanism/benefit
|
Quantitative result (AXT enhancement)
|
Ref
|
| Chemical Mutagenesis + Herbicide |
Improved industrial traits; enhanced growth and pigment accumulation |
30% higher productivity than wild-type (in 25,000-L outdoor raceway ponds). |
105
|
| Three-stage Strategy: UV, EMS, and DPA (Selective Agent) |
Increased cell dry weight; extended logarithmic growth phase |
1.7-fold yield increase (DPA12-2 mutant) over wild-type strain. |
71
|
|
60
Co-γ irradiation under High CO2 |
Enhanced AXT accumulation capacity under optimized conditions |
AXT yield increased to 46.0 mg/L. |
59
|
| UV Mutagenesis + Sodium Azide (Antioxidant Inducer) |
Increased efficiency of carotenoid synthesis; improved stress tolerance |
88.5% increase in AXT content relative to the wild-type culture. |
102
|
| Random Mutagenesis with UV-C |
Higher proliferation rate; increased biomass accumulation |
45% higher AXT accumulation compared to the wild type. |
109
|
Genetic modification based on CRISPR Cas 9
A key advancement in algal biotechnology is the use of RNA-guided endonucleases for targeted genome editing, enabling substantial genetic modifications in freshwater and marine microalgae via CRISPR-Cas9 systems.110
The CRISPR system comprises two classes and six types, with the Cas9-driven type II system widely used for precise gene knockout, knock-in, and knockdown in algal species.110 In microalgae, especially diatoms, CRISPR alters gene functions to improve industrially relevant traits, such as increased lipid accumulation and biomass. The precision of CRISPR-Cas9 genome engineering in microalgae has been enhanced by various techniques.71,111
CRISPR advancements promise to transform research by utilizing photosynthetic cells as scalable platforms for biofuels, food, and chemicals.71,111 This technology has been applied to species including Chlorella spp., Nannochloropsis spp., Synechococcus sp.,Haematococcus sp., Phaeodactylum tricornutum, Chlamydomonas reinhardtii, and Parachlorella kessleri to boost lipids and AXT.112
Specifically in H. pluvialis, CRISPR-Cas9 enables precise editing to enhance AXT synthesis.112 CRISPR-Cas9 targets genes like β-carotene ketolase, enabling knock-in of carotenoid enzymes, yielding a 26.7% increase in AXT (90 mg/L) via enhanced lycopene-to-AXT pathway flux.85 These enzymes mediate the conversion of β-carotene to AXT through sequential hydroxylation and ketolation, providing a direct metabolic enhancement route. CRISPR-Cas9 has also elevated AXT in other microorganisms; for instance, in Xanthophyllomyces dendrorhous, yields increased 1.4-fold over wild type, and in Yarrowia lipolytica, from 44 ± 1 to 285 ± 19 mg/L.113
Electric stimulation (Electrogenetics)
Electrogenetics integrates electronics and genetics, emerging as a field in biology that uses electrical stimulation to remotely control user-defined genetic elements in engineered cells for specific outputs.114 Electric fields precisely modulate these cells' functions.114 A key feature is the use of affordable, accessible devices (like smartphones or rechargeable batteries) for stimulation.115 It also enables precise engineering via micron-scale electrodes, surpassing other triggers like light.114
Electrical stimulation effectively boosts organic compound production in microorganisms, including electroactive, acetogenic, methanogenic species, and mixed cultures.116 This sustainable method uses renewable electricity at ambient conditions without chemicals, producing products like ethanol, butanol, 3-hydroxypropionic acid, 1,3-propanediol, isopropanol, formic acid, butyric acid, and acetate.16,116 However, microalgal studies are limited, with electric treatment outcomes varying by species and cell state.16,116
Kim et al47 exposed biflagellate H. pluvialis cells to 100 mA for 1 min, increasing cell density by 20% but not AXT content after 7 days of photosynthetic cultivation.47 Conversely, Fitriana et al57 applied 30 mA continuously for 2 days, yielding a 36.9% AXT increase (6.0 mg/g cell) over controls.57 These studies demonstrate that prolonged low-intensity stimulation (e.g., 30 mA for 2 days) is more effective than short high-intensity (100 mA for 1 min), yielding 36.9% higher AXT without toxicity.47,57
In aerobic electrochemical conditions, ROS production signals stress response, promoting AXT biosynthesis.15 Excessive treatment damages cells, detaching walls from cytoplasm and causing leaks due to excess radicals and pH drops from electrolysis.57 Electric stimulation enhances cellular redox potential, leading to controlled ROS generation that activates transcription of bkt and crtR -b genes in the carotenoid biosynthetic pathway. This effect promotes the conversion of β-carotene to AXT while simultaneously increasing lipid membrane stability (Fig. 5).16 Fig. 5 demonstrates the pathway from electric stimulation to AXT biosynthesis, aligning with recent findings on bioelectronic control and oxidative stress modulation.114,115,117
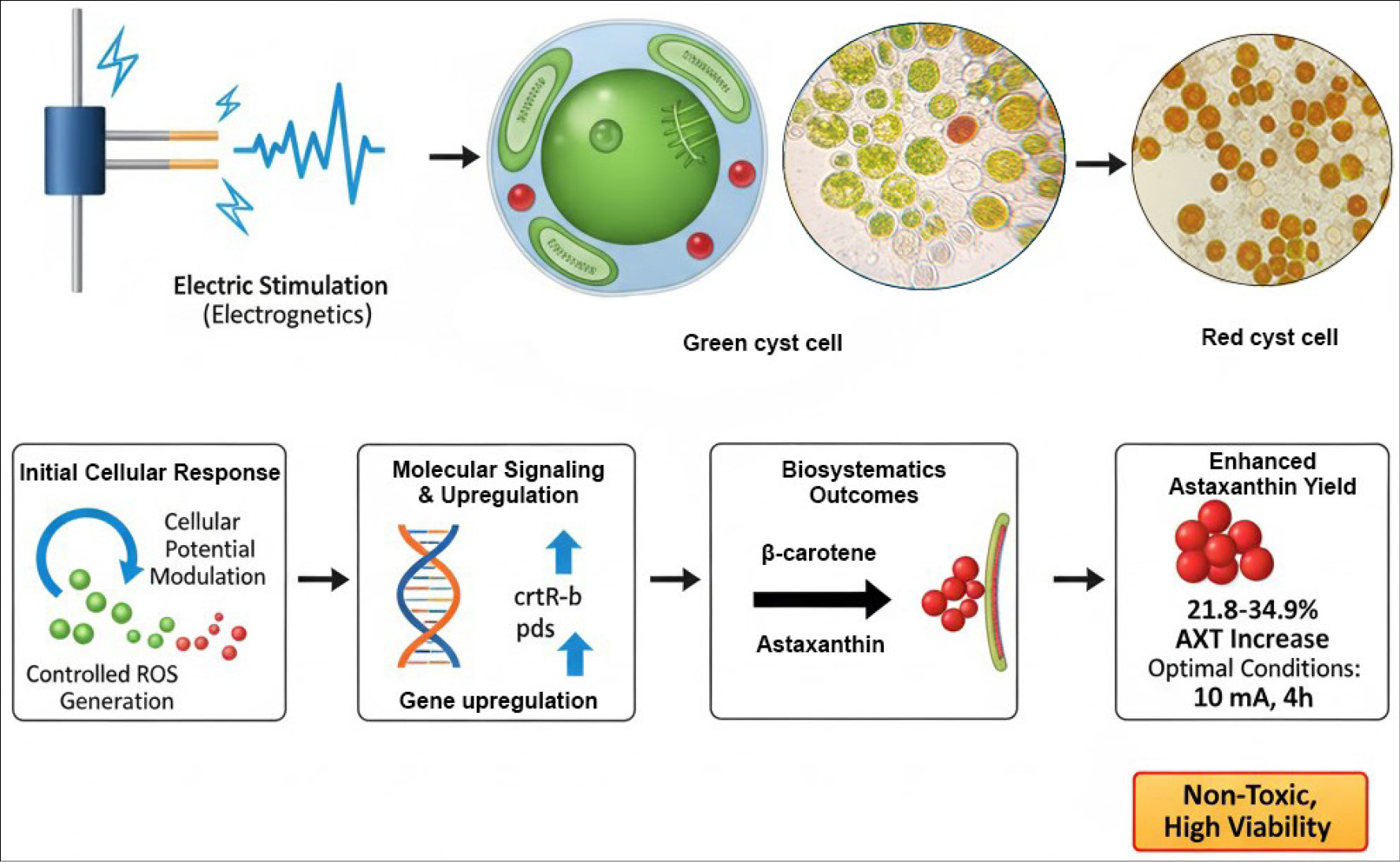
Fig. 5.
Schematic of electric stimulation (electrogenetics) on H. pluvialis, transitioning from green cyst cells to red cyst cells via cellular potential modulation, controlled ROS generation, gene upregulation (e.g., crtR -b, pds), β-carotene to astaxanthin conversion, and enhanced yield (21.8-34.9% AXT increase under optimal conditions: 10 mA, 4h). The process is non-toxic with high viability (based on electrogenetic principles in Refs. 114,115,117).
.
Schematic of electric stimulation (electrogenetics) on H. pluvialis, transitioning from green cyst cells to red cyst cells via cellular potential modulation, controlled ROS generation, gene upregulation (e.g., crtR -b, pds), β-carotene to astaxanthin conversion, and enhanced yield (21.8-34.9% AXT increase under optimal conditions: 10 mA, 4h). The process is non-toxic with high viability (based on electrogenetic principles in Refs. 114,115,117).
In 2023, Sathiyavahisan et al16 identified non-toxic optimal conditions: 10 mA for 4 h rapidly induced AXT in H. pluvialis palmella phase by upregulating pds and crtR -b genes, increasing levels by 21.8-34.9% over controls.16 Prolonged or intense treatments (20–30 mA) reduced AXT and other carotenoids (canthaxanthin, zeaxanthin, β-carotene), lowering viability to 84.2% and 65.6% with morphological changes like faded pigmentation and wall damage.16 Elevated ROS in treated cells likely drives AXT enhancement if treatment is mild.16
Combined strategies to enhance biomass and AXT accumulation
While individual optimization of physical (light, temperature) or chemical (nutrient starvation, CO2 concentration) factors significantly enhances AXT productivity in H. pluvialis, the most advanced and commercially viable strategies often rely on combined stress application to leverage synergistic effects. These multifaceted approaches aim to simultaneously maximize biomass proliferation during the initial green phase and induce rapid encystment and AXT accumulation during the subsequent red phase, sometimes integrating both steps into a continuous process. A highly effective and frequently studied combined strategy is the simultaneous imposition of nitrogen deprivation and high light intensity.58,117 Combined stresses, such as nitrogen deprivation with high light (600 μmol m-2 s-1), yield synergistic effects, increasing AXT by 1.7-fold (174 mg/L) via enhanced ROS signaling and metabolic redirection.46,118,119
Nitrogen starvation halts cell division and photosynthetic machinery repair, while high light stress leads to the excessive generation of ROS. AXT, as a potent antioxidant, is rapidly synthesized within the cell’s cytoplasm and stored in lipid vesicles as a defense mechanism against this oxidative damage, thereby transitioning the cell into the dormant aplanospore stage.117 Studies have demonstrated that this combination results in superior AXT content and productivity compared to sequential or single-factor stress application.58
Beyond the foundational light and nutrient stresses, combining chemical factors and genetic tools has led to notable productivity breakthroughs. For instance, mixotrophic cultivation utilizing organic carbon sources combined with salinity stress has proven effective. Sarada et al explored the combined influence of high salinity and acetate (as an organic carbon source) on H. pluvialis, demonstrating a significant 45% boost in AXT content compared to individual treatments, by providing both stress induction and precursor/energy for biosynthesis.52
Another cutting-edge combined approach integrates genetic or process enhancements with environmental stress. Gómez et al successfully utilized a combination of mutagenesis and nanoparticles for targeted genetic improvement and enhanced induction in a Chilean H. pluvialis strain.105 This sophisticated strategy resulted in a 30% productivity gain of AXT at the mass culture scale, highlighting the potential of merging molecular biology tools with process engineering for hyper-producing high-value carotenoids. Furthermore, combining nutrient limitation with the introduction of redox-sensitive or oxidative stress-inducing compounds has been shown to modulate carotenogenesis, accelerating the accumulation of AXT (Fig. 6).120 As illustrated in Figure 6, combining CRISPR-Cas9 with stress induction has yielded significant enhancements, as reported in recent genetic engineering studies.83,84,101
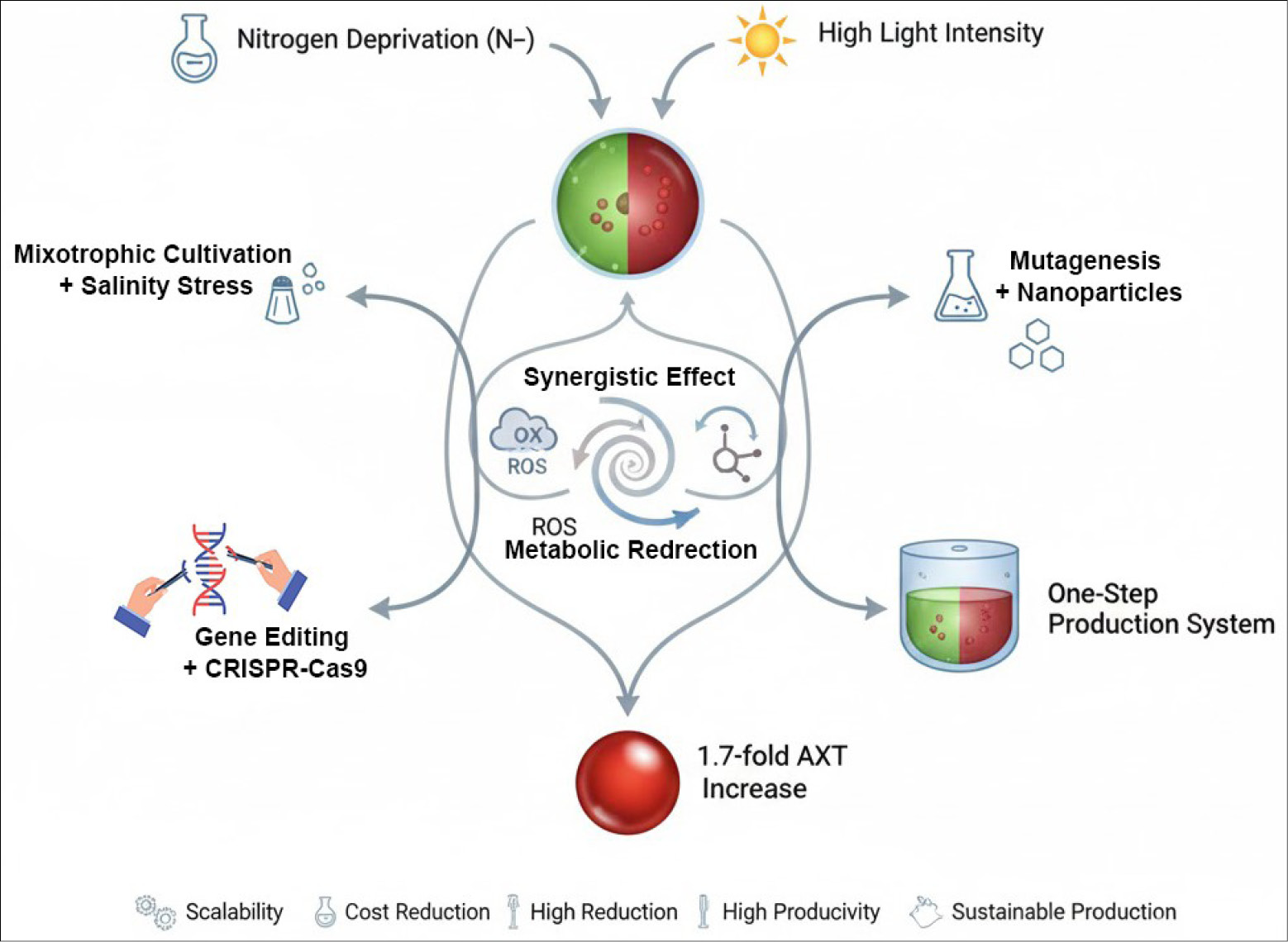
Fig. 6.
Schematic overview of synergistic effects from environmental stressors (nitrogen deprivation, high light intensity, mixotrophic cultivation + salinity stress) and advanced interventions (mutagenesis + nanoparticles, gene editing + CRISPR-Cas9) leading to ROS-mediated metabolic redirection and a 1.7-fold increase in astaxanthin production in a one-step system. Benefits include scalability, cost reduction, high productivity, and sustainable production (inspired by strategies in Refs. 11,83,84,101).
.
Schematic overview of synergistic effects from environmental stressors (nitrogen deprivation, high light intensity, mixotrophic cultivation + salinity stress) and advanced interventions (mutagenesis + nanoparticles, gene editing + CRISPR-Cas9) leading to ROS-mediated metabolic redirection and a 1.7-fold increase in astaxanthin production in a one-step system. Benefits include scalability, cost reduction, high productivity, and sustainable production (inspired by strategies in Refs. 11,83,84,101).
Another pivotal combined approach is the development of one-step production systems.121 This strategy involves continuous culture under nitrogen-limited conditions, where the media is balanced to sustain minimal cell growth (biomass production) while perpetually maintaining the stress level necessary to trigger AXT biosynthesis. This method, successfully tested in various photobioreactors, optimizes resource utilization by eliminating the labor-intensive two-stage process and achieving simultaneous biomass and pigment accumulation.121
The efficacy of such combined and multifaceted approaches underscores the shift in industrial biotechnology towards integrated, continuous, and resource-efficient cultivation systems for natural AXT production.
Conclusion
The escalating global demand for natural, high-value nutraceuticals has positioned microalgae, particularly H. pluvialis, at the forefront of industrial biotechnology. As a consequence, considerable research efforts have been directed towards overcoming the inherent biological and engineering bottlenecks that currently limit the competitive market position of natural AXT against its synthetic counterpart.4,49,122
The comprehensive evaluation of both traditional and emerging biotechnological approaches reviewed herein solidifies a crucial paradigm shift; the future of commercially competitive AXT production from H. pluvialis is intrinsically tied to integrated, multi-stage, and intelligently controlled bioprocessing. While single-factor optimization has been foundational, the industry's profitability hinges on surpassing current volumetric productivity (mg L-1 day-1) limitations to effectively compete against the synthetic market. This endeavor requires transcending empirical protocols through robust synthesis and the adoption of cutting-edge technologies.
The current bottlenecks (low biomass density and prolonged induction time) mandate the implementation of targeted, integrated protocols. This review strongly advocates for the widespread adoption of hybrid cultivation systems, utilizing the economic scalability of open ponds for the initial high-volume green phase and the precision of closed photobioreactors (PBRs) for the AXT-inducing red phase. The critical factor for success in these integrated systems is achieving a final volumetric productivity target of ≥ 25 mg L-1 day-1, which is currently only possible through the synergistic application of precise light quality control (e.g., LED arrays) and optimized chemical stressors. Furthermore, process control must leverage the power of mixotrophy, where the strategic pulse feeding of organic carbon sources, such as acetate, not only fuels growth but also provides essential precursors for the final carotenoid biosynthesis.
To ensure sustained market growth and high-impact scientific contribution, future research must pivot towards quantitative breakthroughs guided by defined metrics. The next generation of AXT hyper-production must be centered on the molecular level: 1) Advanced strain engineering: The utilization of powerful genetic tools, specifically CRISPR-Cas9 and transcriptional engineering, is paramount. The primary research direction should focus on manipulating key regulatory elements (transcription factors) to decouple growth and induction, thereby accelerating the cell cycle. The measurable metric for validating success must be the isolation of a super-strain capable of demonstrating a minimum cellular AXT content of ≥ 5% of dry cell weight combined with a > 20% increase in specific growth rate compared to the current industrial wild-type strains. 2) Smart bioprocess modeling: Industrial control needs to shift from reactive monitoring to predictive automation. Research into advanced machine learning algorithms, such as long short-term memory neural networks, must be pursued to accurately model and forecast AXT yield based on real-time sensor data (pH, ROS, light spectrum). This will enable automated decision-making for optimal stress initiation, with the scientific benchmark for these models set at a predictive accuracy (R2) of ≥ 0.95 for the final product yield.
In conclusion, the trajectory for AXT hyper-production is clear; the integration of biotechnological advances with rigorous process engineering and data-driven intelligence will be the driving force that secures the commercial dominance of this high-value natural AXT.
Review Highlights
What is the current knowledge?
-
Haematococcus pluvialis is the leading natural source for astaxanthin, a potent antioxidant used in nutraceuticals and aquaculture.
-
Two-phase cultivation (green for biomass, red for stress-induced astaxanthin) is standard but faces high costs and low yields.
What is new here?
-
Comprehensive review of emerging biotechnologies like electrogenetics, CRISPR-Cas9, and nanoparticles for hyper-production.
-
Evaluation of combined strategies and quantitative comparisons to guide commercial optimization and reduce costs.
Competing Interests
The authors have no relevant financial or non-financial interests to disclose.
Data Availability Statement
The data underlying this article will be shared on reasonable request to the corresponding author.
Ethical Approval
Not applicable.
References
- Kolašinac SM, Dajić-Stevanović Z, Kilibarda SN, Kostić AŽ. Carotenoids: New applications of “old” pigments. Phyton 2021; 90:1041-62. doi: 10.32604/phyton.2021.015996 [Crossref] [ Google Scholar]
- Maoka T. Carotenoids as natural functional pigments. J Nat Med 2020; 74:1-16. doi: 10.1007/s11418-019-01364-x [Crossref] [ Google Scholar]
- Yuan Z, Zhang Y, Zhang X, Wang L, Ling G, Zhang P. Microalgae and bioactive substances therein: unveiling therapeutic promise against inflammatory bowel disease. J Agric Food Chem 2025; 73:18497-510. doi: 10.1021/acs.jafc.5c04341 [Crossref] [ Google Scholar]
-
Huang JJ, Xie Q, Lin S, Xu W, Cheung PCK. Microalgae-derived astaxanthin: Bioactivities, biotechnological approaches and industrial technologies for its production. Crit Rev Food Sci Nutr 2025; 1-35. doi: 10.1080/10408398.2025.2468863.
- Suyono EA, Pradani L, Mu’avatun U, Habiba RN, Rohma EF. Combination of blue, red, white, and ultraviolet lights for increasing carotenoids and biomass of microalga Haematococcuspluvialis. Procedia Environ Sci 2015; 28:399-405. doi: 10.1016/j.proenv.2015.07.049 [Crossref] [ Google Scholar]
- Garcia-Caparros P, De Filippis L, Gul A, Hasanuzzaman M, Ozturk M, Altay V. Oxidative stress and antioxidant metabolism under adverse environmental conditions: a review. Bot Rev 2021; 87:421-66. doi: 10.1007/s12229-020-09231-1 [Crossref] [ Google Scholar]
- Gong M, Bassi A. Carotenoids from microalgae: A review of recent developments. Biotechnol Adv 2016; 34:1396-412. doi: 10.1016/j.biotechadv.2016.10.005 [Crossref] [ Google Scholar]
- Li X, Wang X, Duan C, Yi S, Gao Z, Xiao C. Biotechnological production of astaxanthin from the microalga Haematococcuspluvialis. Bot Rev 2020; 43:107602. doi: 10.1016/j.biotechadv.2020.107602 [Crossref] [ Google Scholar]
- Mota GCP, Moraes LBSd, Oliveira CYB, Oliveira DWS, Abreu JLd, Dantas DMM. Astaxanthin from Haematococcuspluvialis: processes, applications, and market. Prep BiochemBiotechnol 2022; 52:598-609. doi: 10.1080/10826068.2021.1966802 [Crossref] [ Google Scholar]
- Shah MMR, Liang Y, Cheng JJ, Daroch M. Astaxanthin-producing green microalga Haematococcuspluvialis: from single cell to high value commercial products. Front Plant Sci 2016; 7:531. doi: 10.3389/fpls.2016.00531 [Crossref] [ Google Scholar]
- Patel AK, Tambat VS, Chen C-W, Chauhan AS, Kumar P, Vadrale AP. Recent advancements in astaxanthin production from microalgae: A review. Bioresour Technol 2022; 364:128030. doi: 10.1016/j.biortech.2022.128030 [Crossref] [ Google Scholar]
- Ambati RR, Phang S-M, Ravi S, Aswathanarayana RG. Astaxanthin: Sources, extraction, stability, biological activities and its commercial applications—A review. Mar Drugs 2014; 12:128-52. doi: 10.3390/md12010128 [Crossref] [ Google Scholar]
- Mularczyk M, Michalak I, Marycz K. Astaxanthin and other nutrients from Haematococcuspluvialis—Multifunctional applications. Mar Drugs 2020; 18:459. doi: 10.3390/md18090459 [Crossref] [ Google Scholar]
-
Yusoff FM, Banerjee S, Nagao N, Imaizumi Y, Shariff M, Toda T. Use of microalgae pigments in aquaculture. In: Pigments From Microalgae Handbook. Springer; 2020; 471-513. doi: 10.1007/978-3-030-50971-2_19.
- Li Q, Li L, Zhang Y, Gao H, Zhao Y, Yu X. Chemical inducers regulate ROS signalling to stimulate astaxanthin production in Haematococcuspluvialis under environmental stresses: A review. Trends Food Scie Technol 2023; 136:181-93. doi: 10.1016/j.tifs.2023.04.014 [Crossref] [ Google Scholar]
- Sathiyavahisan LP, Lakshmi Narasimhan A, Mahadi R, Kim S, Christabel C, Yu H. Rapid induction of astaxanthin in Haematococcuslacustris by mild electric stimulation. Appl Sci 2023; 13:12959. doi: 10.3390/app132312959 [Crossref] [ Google Scholar]
- Farruggia C, Kim M-B, Bae M, Lee Y, Pham TX, Yang Y. Astaxanthin exerts anti-inflammatory and antioxidant effects in macrophages in NRF2-dependent and independent manners. J NutrBiochem 2018; 62:202-9. doi: 10.1016/j.jnutbio.2018.09.005 [Crossref] [ Google Scholar]
- Wang B, Pan X, Jia J, Xiong W, Manirafasha E, Ling X. Strategy and regulatory mechanisms of glutamate feeding to enhance astaxanthin yield in Xanthophyllomyces dendrorhous. Enzyme Microb Technol 2019; 125:45-52. doi: 10.1016/j.enzmictec.2019.02.010 [Crossref] [ Google Scholar]
- Kim B, Lee SY, Narasimhan AL, Kim S, Oh Y-K. Cell disruption and astaxanthin extraction from Haematococcuspluvialis: Recent advances. Bioresour Technol 2022; 343:126124. doi: 10.1016/j.biortech.2021.126124 [Crossref] [ Google Scholar]
- Kang Y, Xu L, Dong J, Yuan X, Ye J, Fan Y. Programmed microalgae-gel promotes chronic wound healing in diabetes. Nat Commun 2024; 15:1042. doi: 10.1038/s41467-024-45101-9 [Crossref] [ Google Scholar]
- Niizawa I, Espinaco BY, Leonardi JR, Heinrich JM, Sihufe GA. Enhancement of astaxanthin production from Haematococcuspluvialis under autotrophic growth conditions by a sequential stress strategy. Prep BiochemBiotechnol 2018; 48:528-34. doi: 10.1080/10826068.2018.1466159 [Crossref] [ Google Scholar]
- Zhao T, Yan X, Sun L, Yang T, Hu X, He Z. Research progress on extraction, biological activities and delivery systems of natural astaxanthin. Trends Food Sci Technol 2019; 91:354-61. doi: 10.1016/j.tifs.2019.07.014 [Crossref] [ Google Scholar]
- Han S-I, Chang SH, Lee C, Jeon MS, Heo YM, Kim S. Astaxanthin biosynthesis promotion with pH shock in the green microalga, Haematococcuslacustris. Bioresour Technol 2020; 314:123725. doi: 10.1016/j.biortech.2020.123725 [Crossref] [ Google Scholar]
- Brendler T, Williamson EM. Astaxanthin: How much is too much? A safety review. Phytother Res 2019; 33:3090-111. doi: 10.1002/ptr.6514 [Crossref] [ Google Scholar]
- Capelli B, Bagchi D, Cysewski GR. Synthetic astaxanthin is significantly inferior to algal-based astaxanthin as an antioxidant and may not be suitable as a human nutraceutical supplement. Nutrafoods 2013; 12:145-52. doi: 10.1007/s13749-013-0051-5 [Crossref] [ Google Scholar]
-
Tambat VS, Singhania RR, Sumathi Y, Chen C-W, Dong C-D, Michaud P, et al. Astaxanthin: nature’s multifunctional molecule, natural sources, health benefits, and process advancements. Crit Rev Biotechnol 2025; 1-19. doi: 10.1080/07388551.2025.2537816.
- Mao X, Wu T, Sun D, Zhang Z, Chen F. Differential responses of the green microalga Chlorella zofingiensis to the starvation of various nutrients for oil and astaxanthin production. Bioresour Technol 2018; 249:791-8. doi: 10.1016/j.biortech.2017.10.090 [Crossref] [ Google Scholar]
- Tramontin LRR, Kildegaard KR, Sudarsan S, Borodina I. Enhancement of astaxanthin biosynthesis in oleaginous yeast Yarrowia lipolytica via microalgal pathway. Microorganisms 2019; 7:472. doi: 10.3390/microorganisms7100472 [Crossref] [ Google Scholar]
- Ha PT, Hoang NH, Lien NTK, Phuong NTD, Huy NQ, Thoa NK. Selection of bacterial strains belonging to the astaxanthin producing Paracoccus genus. Vietnam J Biotechnol 2018; 16:565-72. doi: 10.15625/1811-4989/16/3/13477 [Crossref] [ Google Scholar]
- Oslan SNH, Shoparwe NF, Yusoff AH, Rahim AA, Chang CS, Tan JS. A review on Haematococcuspluvialis bioprocess optimization of green and red stage culture conditions for the production of natural astaxanthin. Biomolecules 2021; 11:256. doi: 10.3390/biom11020256 [Crossref] [ Google Scholar]
- Watanabe K, Arafiles KHV, Higashi R, Okamura Y, Tajima T, Matsumura Y. Isolation of high carotenoid-producing Aurantiochytrium sp mutants and improvement of astaxanthin productivity using metabolic information. J Oleo Sci 2018; 67:571-8. doi: 10.5650/jos.ess17230 [Crossref] [ Google Scholar]
- Lim KC, Yusoff FM, Shariff M, Kamarudin MS. Astaxanthin as feed supplement in aquatic animals. Rev Aquac 2018; 10:738-73. doi: 10.1111/raq.12200 [Crossref] [ Google Scholar]
- Cheong JY, Muskhazli M, Ahmad SA, Adli AA. Three dimensional optimisation for the enhancement of astaxanthin recovery from shrimp shell wastes by Aeromonas hydrophila. Biocatal Agric Biotechnol 2020; 27:101649. doi: 10.1016/j.bcab.2020.101649 [Crossref] [ Google Scholar]
- Azizi M, Moteshafi H, Hashemi M. Distinctive nutrient designs using statistical approach coupled with light feeding strategy to improve the Haematococcuspluvialis growth performance and astaxanthin accumulation. Bioresour Technol 2020; 300:122594. doi: 10.1016/j.biortech.2019.122594 [Crossref] [ Google Scholar]
- Hong ME, Chang WS, Patel AK, Oh MS, Lee JJ, Sim SJ. Microalgal-based carbon sequestration by converting LNG-fired waste CO2 into red gold astaxanthin: the potential applicability. Energies 2019; 12:1718. doi: 10.3390/en12091718 [Crossref] [ Google Scholar]
- Wang F, Gao B, Wu M, Huang L, Zhang C. A novel strategy for the hyper-production of astaxanthin from the newly isolated microalga Haematococcuspluvialis JNU35. Algal Res 2019; 39:101466. doi: 10.1016/j.algal.2019.101466 [Crossref] [ Google Scholar]
- Tang S, Liu Y, Zhu J, Cheng X, Liu L, Hammerschmidt K. Bet hedging in a unicellular microalga. Nat Commun 2024; 15:2063. doi: 10.1038/s41467-024-46297-6 [Crossref] [ Google Scholar]
- Zhang C, Liu J, Zhang L. Cell cycles and proliferation patterns in Haematococcuspluvialis. Chin J OceanolLimnol 2017; 35:1205-11. doi: 10.1007/s00343-017-6103-8 [Crossref] [ Google Scholar]
- Fábregas J, Otero A, Maseda A, Domínguez A. Two-stage cultures for the production of astaxanthin from Haematococcuspluvialis. J Biotechnol 2001; 89:65-71. doi: 10.1016/S0168-1656(01)00289-9 [Crossref] [ Google Scholar]
-
Han D, Li Y, Hu Q. Biology and commercial aspects of Haematococcuspluvialis. In: Handbook of Microalgal Culture: Applied Phycology and Biotechnology. Wiley; 2013. p. 388-405. doi: 10.1002/9781118567166.ch20.
-
Liu J, van der Meer JP, Zhang L, Zhang Y. 12. Cultivation of Haematococcuspluvialis for astaxanthin production. In: Microalgal Production for Biomass and High‐Value Products. CRC Press; 2017. p. 267-93. doi: 10.1201/b19464.
- Gherabli A, Grimi N, Lemaire J, Vorobiev E, Lebovka N. Extraction of valuable biomolecules from the microalga Haematococcuspluvialis assisted by electrotechnologies. Molecules 2023; 28:2089. doi: 10.3390/molecules28052089 [Crossref] [ Google Scholar]
- Nemani N, Dehnavi SM, Pazuki G. Extraction and separation of astaxanthin with the help of pre-treatment of Haematococcuspluvialis microalgae biomass using aqueous two-phase systems based on deep eutectic solvents. Sci Rep 2024; 14:5420. doi: 10.1038/s41598-024-55630-4 [Crossref] [ Google Scholar]
- Doria E, Temporiti MEE, Damiani MC, Popovich CA, Leonardi PI, Nielsen E. Influence of light stress on the accumulation of xanthophylls and lipids in Haematococcuspluvialis CCALA 1081 grown under autotrophic or mixotrophic conditions. Journal of Marine Biology and Aquaculture 2018; 4:30-35. [ Google Scholar]
- Pang N, Chen S. Effects of C5 organic carbon and light on growth and cell activity of Haematococcuspluvialis under mixotrophic conditions. Algal Res 2017; 21:227-35. doi: 10.1016/j.algal.2016.12.003 [Crossref] [ Google Scholar]
- Christian D, Zhang J, Sawdon AJ, Peng C-A. Enhanced astaxanthin accumulation in Haematococcuspluvialis using high carbon dioxide concentration and light illumination. Bioresour Technol 2018; 256:548-51. doi: 10.1016/j.biortech.2018.02.074 [Crossref] [ Google Scholar]
- Kim JY, Lee C, Jeon MS, Park J, Choi Y-E. Enhancement of microalga Haematococcuspluvialis growth and astaxanthin production by electrical treatment. Bioresour Technol 2018; 268:815-9. doi: 10.1016/j.biortech.2018.08.014 [Crossref] [ Google Scholar]
- Cheng J, Li K, Yang Z, Zhou J, Cen K. Enhancing the growth rate and astaxanthin yield of Haematococcuspluvialis by nuclear irradiation and high concentration of carbon dioxide stress. Bioresour Technol 2016; 204:49-54. doi: 10.1016/j.biortech.2015.12.076 [Crossref] [ Google Scholar]
- Cui H, Zhu X, Yu X, Li S, Wang K, Wei L. Advancements of astaxanthin production in Haematococcuspluvialis: Update insight and way forward. Biotechnol Adv 2025; 79:108519. doi: 10.1016/j.biotechadv.2025.108519 [Crossref] [ Google Scholar]
- Patel AK, Joun JM, Hong ME, Sim SJ. Effect of light conditions on mixotrophic cultivation of green microalgae. Bioresour Technol 2019; 282:245-53. doi: 10.1016/j.biortech.2019.03.024 [Crossref] [ Google Scholar]
- Pereira S, Otero A. Haematococcuspluvialis bioprocess optimization: Effect of light quality, temperature and irradiance on growth, pigment content and photosynthetic response. Algal Res 2020; 51:102027. doi: 10.1016/j.algal.2020.102027 [Crossref] [ Google Scholar]
- Sarada R, Tripathi U, Ravishankar G. Influence of stress on astaxanthin production in Haematococcuspluvialis grown under different culture conditions. Process Biochem 2002; 37:623-7. doi: 10.1016/S0032-9592(01)00246-1 [Crossref] [ Google Scholar]
- Tripathi U, Sarada R, Ravishankar G. Effect of culture conditions on growth of green alga—Haematococcuspluvialis and astaxanthin production. Acta Physiol Plant 2002; 24:323-9. doi: 10.1007/s11738-002-0058-9 [Crossref] [ Google Scholar]
- Tam LT, Hoang DD, Mai DTN, Thu NTH, Anh HTL, Hong DD. Study on the effect of salt concentration on growth and Astaxanthin accumulation of microalgae Haematococcuspluvialis as the initial basis for two phase culture of astaxanthin production. Acad J Biol 2012; 34:213-23. doi: 10.15625/0866-7160/v34n2.964 [Crossref] [ Google Scholar]
- Gao Z, Meng C, Chen YC, Ahmed F, Mangott A, Schenk PM. Comparison of astaxanthin accumulation and biosynthesis gene expression of three Haematococcuspluvialis strains upon salinity stress. J Appl Psychol 2015; 27:1853-60. doi: 10.1007/s10811-014-0491-3 [Crossref] [ Google Scholar]
- Hwang S-W, Choi HI, Sim SJ. Acidic cultivation of Haematococcuspluvialis for improved astaxanthin production in the presence of a lethal fungus. Bioresour Technol 2019; 278:138-44. doi: 10.1016/j.biortech.2019.01.080 [Crossref] [ Google Scholar]
- Fitriana H-N, Lee S-Y, Choi S-A, Lee J-Y, Kim B-L, Lee J-S. Electric stimulation of astaxanthin biosynthesis in Haematococcuspluvialis. Appl Sci 2021; 11:3348. doi: 10.3390/app11083348 [Crossref] [ Google Scholar]
- Scibilia L, Girolomoni L, Berteotti S, Alboresi A, Ballottari M. Photosynthetic response to nitrogen starvation and high light in Haematococcuspluvialis. Algal Res 2015; 12:170-81. doi: 10.1016/j.algal.2015.08.024 [Crossref] [ Google Scholar]
- Cheng J, Li K, Yang Z, Lu H, Zhou J, Cen K. Gradient domestication of Haematococcuspluvialis mutant with 15% CO2 to promote biomass growth and astaxanthin yield. Bioresour Technol 2016; 216:340-4. doi: 10.1016/j.biortech.2016.05.095 [Crossref] [ Google Scholar]
- Nguyen KD. Astaxanthin: a comparative case of synthetic vs. natural production. Chemical and Biomolecular Engineering Publications and Other Works 2013. http://trace.tennessee.edu/utk_chembiopubs/94.
- Khoo KS, Lee SY, Ooi CW, Fu X, Miao X, Ling TC. Recent advances in biorefinery of astaxanthin from Haematococcuspluvialis. Bioresour Technol 2019; 288:121606. doi: 10.1016/j.biortech.2019.121606 [Crossref] [ Google Scholar]
- Li C, Gao Y, Huan Y, Ren P, Zhi J, Wu A. Colon and gut microbiota greatly affect the absorption and utilization of astaxanthin derived from Haematococcuspluvialis. Food Res Int 2022; 156:111324. doi: 10.1016/j.foodres.2022.111324 [Crossref] [ Google Scholar]
- Ahuja K, Rawat A. Astaxanthin market size by source (synthetic, natural), by application (dietary supplement, personal care, pharmaceuticals, food and beverages, animal feed) industry outlook report, regional analysis, application potential, price trends, competitive market and forecast, 2019–2026. Global Market Insights, Selbyville, DE, USA; 2019. https://www.marketresearch.com/One-Off-Global-Market-Insights-v4130/Astaxanthin-Size-Source-Synthetic-Natural-14839588. Accessed on 20 January 2022.
- Leu S, Boussiba S. Advances in the production of high-value products by microalgae. Ind Biotechnol 2014; 10:169-83. doi: 10.1089/ind.2013.0039 [Crossref] [ Google Scholar]
- Li J, Zhu D, Niu J, Shen S, Wang G. An economic assessment of astaxanthin production by large scale cultivation of Haematococcuspluvialis. Biotechnol Adv 2011; 29:568-74. doi: 10.1016/j.biotechadv.2011.04.001 [Crossref] [ Google Scholar]
- Pang N, Fu X, Fernandez JSM, Chen S. Multilevel heuristic LED regime for stimulating lipid and bioproducts biosynthesis in Haematococcuspluvialis under mixotrophic conditions. Bioresour Technol 2019; 288:121525. doi: 10.1016/j.biortech.2019.121525 [Crossref] [ Google Scholar]
-
Chen J-H, Wei D, Lim P-E, Xie J, Chen WN. Screening and effect evaluation of chemical inducers for enhancing astaxanthin and lipid production in mixotrophic Chromochloris zofingiensis. J Appl Psychol 2022; 1-18. doi: 10.1007/s10811-021-02618-6.
- Khorshidi N, Hassanpour H, Ziyadi H. Static magnetic field improved growth and astaxanthin production in Haematococcus lacustris via the regulation of carbohydrate accumulation, H2O2 level, and antioxidant defense system. J Appl Psychol 2022; 34:2283-95. doi: 10.1007/s10811-022-02758-3 [Crossref] [ Google Scholar]
- Yu C, Wang H-P, Yu X. The associative induction of succinic acid and hydrogen sulfide for high-producing biomass, astaxanthin and lipids in Haematococcuspluvialis. Bioresour Technol 2022; 358:127397. doi: 10.1016/j.biortech.2022.127397 [Crossref] [ Google Scholar]
- Evens TJ, Niedz RP, Kirkpatrick GJ. Temperature and irradiance impacts on the growth, pigmentation and photosystem II quantum yields of Haematococcuspluvialis (Chlorophyceae). J Appl Psychol 2008; 20:411-22. doi: 10.1007/s10811-007-9277-1 [Crossref] [ Google Scholar]
- Wang N, Guan B, Kong Q, Sun H, Geng Z, Duan L. Enhancement of astaxanthin production from Haematococcuspluvialis mutants by three-stage mutagenesis breeding. J Biotechnol 2016; 236:71-7. doi: 10.1016/j.jbiotec.2016.08.009 [Crossref] [ Google Scholar]
- Zhang W-w, Zhou X-f, Zhang Y-l, Cheng P-f, Ma R, Cheng W-l. Enhancing astaxanthin accumulation in Haematococcuspluvialis by coupled light intensity and nitrogen starvation in column photobioreactors. J Microbiol Biotechnol 2018; 28:2019-28. doi: 10.4014/jmb.1807.07008 [Crossref] [ Google Scholar]
- Xi T, Kim DG, Roh SW, Choi J-S, Choi Y-E. Enhancement of astaxanthin production using Haematococcuspluvialis with novel LED wavelength shift strategy. Appl Microbiol Biotechnol 2016; 100:6231-8. doi: 10.1007/s00253-016-7301-6 [Crossref] [ Google Scholar]
- Lee C, Ahn J-W, Kim J-B, Kim JY, Choi Y-E. Comparative transcriptome analysis of Haematococcuspluvialis on astaxanthin biosynthesis in response to irradiation with red or blue LED wavelength. World J Microbiol Biotechnol 2018; 34:1-14. doi: 10.1007/s11274-018-2459-y [Crossref] [ Google Scholar]
- Giannelli L, Yamada H, Katsuda T, Yamaji H. Effects of temperature on the astaxanthin productivity and light harvesting characteristics of the green alga Haematococcuspluvialis. J BiosciBioeng 2015; 119:345-50. doi: 10.1016/j.jbiosc.2014.09.002 [Crossref] [ Google Scholar]
- Cong X, Zang X, Dong M, Wang Z, He B, Hou L. Accumulation of phytoene and astaxanthin and related genes expression in Haematococcuspluvialis under sodium acetate stress. Aquat Biol 2020; 29:155-64. doi: 10.3354/ab00733 [Crossref] [ Google Scholar]
- Jin C, You J, Zhou Z, Liu Q, Zhou X. Novel insights into saline stress on photosynthetic activity and astaxanthin production of Haematococcuspluvialis. J OceanolLimnol 2025; 43:921-38. doi: 10.1007/s00343-024-4104-y [Crossref] [ Google Scholar]
- Choi YY, Joun JM, Lee J, Hong ME, Pham H-M, Chang WS. Development of large-scale and economic pH control system for outdoor cultivation of microalgae Haematococcuspluvialis using industrial flue gas. Bioresour Technol 2017; 244:1235-44. doi: 10.1016/j.biortech.2017.05.147 [Crossref] [ Google Scholar]
- Nasri N, Keyhanfar M, Behbahani M, Dini G. Enhancement of astaxanthin production in Haematococcuspluvialis using zinc oxide nanoparticles. J Biotechnol 2021; 342:72-8. doi: 10.1016/j.jbiotec.2021.10.004 [Crossref] [ Google Scholar]
- Kim Y-E, Matter IA, Lee N, Jung M, Lee Y-C, Choi S-A. Enhancement of astaxanthin production by Haematococcuspluvialis using magnesium aminoclay nanoparticles. Bioresour Technol 2020; 307:123270. doi: 10.1016/j.biortech.2020.123270 [Crossref] [ Google Scholar]
- Zhang Z, Wang B, Hu Q, Sommerfeld M, Li Y, Han D. A new paradigm for producing astaxanthin from the unicellular green alga Haematococcuspluvialis. BiotechnolBioeng 2016; 113:2088-99. doi: 10.1002/bit.25976 [Crossref] [ Google Scholar]
- Radice RP, Padula MC, Liguori A, D’Arienzo G, Martelli G. Genetic improvement to obtain specialized Haematococcuspluvialis genotypes for the production of carotenoids, with particular reference to astaxanthin. Int J Plant Biol 2023; 14:276-85. doi: 10.3390/ijpb14010023 [Crossref] [ Google Scholar]
- Acheampong A, Li L, Elsherbiny SM, Wu Y, Swallah MS, Bondzie-Quaye P. A crosswalk on the genetic and conventional strategies for enhancing astaxanthin production in Haematococcuspluvialis. Crit Rev Biotechnol 2024; 44:1018-39. doi: 10.1080/07388551.2023.2240009 [Crossref] [ Google Scholar]
- Kayani S-I, -Rahman S-u, Shen Q, Cui Y, Liu W, Hu X. Molecular approaches to enhance astaxanthin biosynthesis; future outlook: engineering of transcription factors in Haematococcuspluvialis. Crit Rev Biotechnol 2024; 44:514-29. doi: 10.1080/07388551.2023.2208284 [Crossref] [ Google Scholar]
- Xie Q, Li S, Zhao D, Ye L, Li Q, Zhang X. Manipulating the position of DNA expression cassettes using location tags fused to dCas9 (Cas9-Lag) to improve metabolic pathway efficiency. Microb Cell Fact 2020; 19:1-11. doi: 10.1186/s12934-020-01496-w [Crossref] [ Google Scholar]
- Domınguez-Bocanegra A, Legarreta IG, Jeronimo FM, Campocosio AT. Influence of environmental and nutritional factors in the production of astaxanthin from Haematococcuspluvialis. Bioresour Technol 2004; 92:209-14. doi: 10.1016/j.biortech.2003.04.001 [Crossref] [ Google Scholar]
- Wong Y. Effects of light intensity, illumination cycles on microalgae Haematococcuspluvialis for production of astaxanthin. J Mar Biol Aquacult 2016; 2:1-6. doi: 10.15436/2381-0750.16.1083 [Crossref] [ Google Scholar]
- Torzillo G, Göksan T, Isik O, Gökpinar Ş. Photon irradiance required to support optimal growth and interrelations between irradiance and pigment composition in the green alga Haematococcuspluvialis. Eur J Phycol 2005; 40:233-40. doi: 10.1080/09670260500123609 [Crossref] [ Google Scholar]
- Lv H, Xia F, Liu M, Cui X, Wahid F, Jia S. Metabolomic profiling of the astaxanthin accumulation process induced by high light in Haematococcuspluvialis. Algal Res 2016; 20:35-43. doi: 10.1016/j.algal.2016.09.019 [Crossref] [ Google Scholar]
- Xi T, Kim DG, Roh SW, Choi J-S, Choi Y-E. Enhancement of astaxanthin production using Haematococcuspluvialis with novel LED wavelength shift strategy. Appl Microbiol Biotechnol 2016; 100:6231-8. doi: 10.1007/s00253-016-7301-6 [Crossref] [ Google Scholar]
- Kim Z-H, Lee H-S, Lee C-G. Red and blue photons can enhance the production of astaxanthin from Haematococcuspluvialis. Algae 2009; 24:121-7. doi: 10.4490/algae.2009.24.2.121 [Crossref] [ Google Scholar]
- Park E-K, Lee C-G. Astaxanthin production by Haematococcuspluvialis under various light intensities and wavelengths. J Microbiol Biotechnol 2001; 11:1024-30. [ Google Scholar]
- Hong M-E, Hwang SK, Chang WS, Kim BW, Lee J, Sim SJ. Enhanced autotrophic astaxanthin production from Haematococcuspluvialis under high temperature via heat stress-driven Haber–Weiss reaction. Appl Microbiol Biotechnol 2015; 99:5203-15. [ Google Scholar]
- Hanan NA, Al-Shorgani NK, Shukor H, Kalil MS. Pre-optimization conditions for Haematococcuspluvialis growth. International Journal on Advanced Science, Engineering and Information Technology 2013; 3:168-71. [ Google Scholar]
- Liyanaarachchi VC, Nishshanka GKSH, Premaratne RGMM, Ariyadasa TU, Nimarshana PHV, Malik A. Astaxanthin accumulation in the green microalga Haematococcuspluvialis: Effect of initial phosphate concentration and stepwise/continuous light stress. Biotechnol Rep 2020; 28:e00538. doi: 10.1016/j.btre.2020.e00538 [Crossref] [ Google Scholar]
- Oslan SNH, Tan JS, Oslan SN, Matanjun P, Mokhtar RAM, Shapawi R. Haematococcuspluvialis as a potential source of astaxanthin with diverse applications in industrial sectors: current research and future directions. Molecules 2021; 26:6470. doi: 10.3390/molecules26216470 [Crossref] [ Google Scholar]
- Sun Y-h, Cui S-g, Zhang Y-l. The Effect of pH on the Accumulation of Astaxanthin in HaematococcusPluvialis in the Induction Stage. Modern Chemistry 2022; 10:23-9. doi: 10.11648/j.mc.20221002.11 [Crossref] [ Google Scholar]
- Choi S-A, Jeong Y, Lee J, Huh YH, Choi SH, Kim H-S. Biocompatible liquid-type carbon nanodots (C-paints) as light delivery materials for cell growth and astaxanthin induction of Haematococcuspluvialis. Mater Sci Eng C 2020; 109:110500. doi: 10.1016/j.msec.2019.110500 [Crossref] [ Google Scholar]
- Hee CW, Shing WL, Tian LM, Yoke CM, Hock OG. Effects of silver nanoparticles on the carotenoid production from Haematococcuspluvialis. EnvironmentAsia 2020; 13:106-111. doi: 10.14456/ea.2020.10 [Crossref] [ Google Scholar]
- Rastar M, Hosseini Shekarabi S, Shamsaie Mehrgan M, Sabzi S. Effects of iron and zinc concentrations on growth performance and biochemical composition of Haematococcuspluvialis: A comparison between nanoparticles and their corresponding metals bulks. J Algal Biomass Util 2018; 9:59-67. [ Google Scholar]
- Galarza JI, Gimpel JA, Rojas V, Arredondo-Vega BO, Henríquez V. Over-accumulation of astaxanthin in Haematococcuspluvialis through chloroplast genetic engineering. Algal Res 2018; 31:291-7. doi: 10.1016/j.algal.2018.02.024 [Crossref] [ Google Scholar]
- Hong ME, Choi HI, Kwak HS, Hwang S-W, Sung YJ, Chang WS. Rapid selection of astaxanthin-hyperproducing Haematococcus mutant via azide-based colorimetric assay combined with oil-based astaxanthin extraction. Bioresour Technol 2018; 267:175-81. doi: 10.1016/j.biortech.2018.07.033 [Crossref] [ Google Scholar]
- Trovão M, Schüler LM, Machado A, Bombo G, Navalho S, Barros A. Random mutagenesis as a promising tool for microalgal strain improvement towards industrial production. Mar Drugs 2022; 20:440. doi: 10.3390/md20070440 [Crossref] [ Google Scholar]
- Hu Z, Li Y, Sommerfeld M, Chen F, Hu Q. Enhanced protection against oxidative stress in an astaxanthin-overproduction Haematococcus mutant (Chlorophyceae). Eur J Phycol 2008; 43:365-76. doi: 10.1080/09670260802227736 [Crossref] [ Google Scholar]
- Gómez PI, Inostroza I, Pizarro M, Pérez J. From genetic improvement to commercial-scale mass culture of a Chilean strain of the green microalga Haematococcuspluvialis with enhanced productivity of the red ketocarotenoid astaxanthin. AoB Plants 2013; 5:plt026. doi: 10.1093/aobpla/plt026 [Crossref] [ Google Scholar]
- Tripathi U, Venkateshwaran G, Sarada R, Ravishankar G. Studies on Haematococcuspluvialis for improved production of astaxanthin by mutagenesis. World J Microbiol Biotechnol 2001; 17:143-8. doi: 10.1023/A:1016609815405 [Crossref] [ Google Scholar]
- Chen Y, Li D, Lu W, Xing J, Hui B, Han Y. Screening and characterization of astaxanthin-hyperproducing mutants of Haematococcuspluvialis. Biotechnol Lett 2003; 25:527-9. doi: 10.1023/A:1022877703008 [Crossref] [ Google Scholar]
- Liu J, Huang Q. Screening of astaxanthin-hyperproducing Haematococcuspluvialis using Fourier transform infrared (FT-IR) and Raman microspectroscopy. Appl Spectrosc 2016; 70:1639-48. doi: 10.1177/0003702816645605 [Crossref] [ Google Scholar]
- Telli M, Ünlü ES. Comparative de novo transcriptome analysis and random UV mudtagenesis: application in high biomass and astaxanthin production enhancement for Haematococcuspluvialis. Mol Biol Rep 2023; 50:8133-43. doi: 10.1007/s11033-023-08722-9 [Crossref] [ Google Scholar]
- Patel VK, Soni N, Prasad V, Sapre A, Dasgupta S, Bhadra B. CRISPR–Cas9 system for genome engineering of photosynthetic microalgae. Mol Biotechnol 2019; 61:541-61. doi: 10.1007/s12033-019-00185-3 [Crossref] [ Google Scholar]
- Wang Q, Lu Y, Xin Y, Wei L, Huang S, Xu J. Genome editing of model oleaginous microalgae Nannochloropsis spp by CRISPR/Cas9. Plant J 2016; 88:1071-81. doi: 10.1111/tpj.13307 [Crossref] [ Google Scholar]
- Le-Feuvre R, Moraga-Suazo P, Gonzalez J, Martin SS, Henríquez V, Donoso A. Biotechnology applied to Haematococcuspluvialis Fotow: challenges and prospects for the enhancement of astaxanthin accumulation. J Appl Psychol 2020; 32:3831-52. doi: 10.1007/s10811-020-02231-z [Crossref] [ Google Scholar]
- Yamamoto K, Hara KY, Morita T, Nishimura A, Sasaki D, Ishii J. Enhancement of astaxanthin production in Xanthophyllomyces dendrorhous by efficient method for the complete deletion of genes. Microb Cell Fact 2016; 15:1-8. doi: 10.1186/s12934-016-0556-x [Crossref] [ Google Scholar]
- Mansouri M, Fussenegger M. Electrogenetics: bridging synthetic biology and electronics to remotely control the behavior of mammalian designer cells. CurrOpin Chem Biol 2022; 68:102151. doi: 10.1016/j.cbpa.2022.102151 [Crossref] [ Google Scholar]
- Terrell JL, Tschirhart T, Jahnke JP, Stephens K, Liu Y, Dong H. Bioelectronic control of a microbial community using surface-assembled electrogenetic cells to route signals. Nat Nanotechnol 2021; 16:688-97. doi: 10.1038/s41565-021-00878-4 [Crossref] [ Google Scholar]
- Singh NK, Mathuriya AS, Mehrotra S, Pandit S, Singh A, Jadhav D. Advances in bioelectrochemical systems for bio-products recovery. Environ Technol 2024; 45:3853. doi: 10.1080/09593330.2023.2234676 [Crossref] [ Google Scholar]
- Cray R, Levine I. Oxidative stress modulates astaxanthin synthesis in Haematococcuspluvialis. J Appl Psychol 2022; 34:2327-38. doi: 10.1007/s10811-022-02792-1 [Crossref] [ Google Scholar]
- Zhang L, Zhang C, Xu R, Yu W, Liu J. A strategy for promoting carbon flux into fatty acid and astaxanthin biosynthesis by inhibiting the alternative oxidase respiratory pathway in Haematococcuspluvialis. Bioresour Technol 2022; 344:126275. doi: 10.1016/j.biortech.2021.126275 [Crossref] [ Google Scholar]
- González-Delgado AD, Torres LK, SUAREZ GELVEZ JH, Urbina-Suarez NA. Astaxanthin production from Haematococcuspluvialis: Effects of light wavelength and salinity. Contemporary Engineering Sciences 2017; 10:1739-46. doi: 10.12988/ces.2017.711196 [Crossref] [ Google Scholar]
- Sung YJ, Sim SJ. Multifaceted strategies for economic production of microalgae Haematococcuspluvialis-derived astaxanthin via direct conversion of CO2. Bioresour Technol 2022; 344:126255. doi: 10.1016/j.biortech.2021.126255 [Crossref] [ Google Scholar]
- Del Río E, Acién FG, García‐Malea MC, Rivas J, Molina‐Grima E, Guerrero MG. Efficiency assessment of the one‐step production of astaxanthin by the microalga Haematococcuspluvialis. BiotechnolBioeng 2008; 100:397-402. doi: 10.1002/bit.21770 [Crossref] [ Google Scholar]
- Jaryal S, Sharma AK, Yadav Lamba B, PATEL A. A Comprehensive Review on Microalgae based Astaxanthin: Bioprocess Optimization, Technological Barriers, Industrial Applications and Future Roadmap. Front Mar Sci 2025; 12:1644644. doi: 10.3389/fmars.2025.1644644 [Crossref] [ Google Scholar]