Bioimpacts. 16:32772.
doi: 10.34172/bi.32772
Original Article
Enhancing cisplatin chemosensitivity in gastric cancer through LINC00162 silencing: Modulation of the PI3K/AKT pathway and NANOG downregulation
Seyed Ali Hosseini Conceptualization, Data curation, Funding acquisition, Investigation, Software, Visualization, Writing – original draft, 1 
Amir Ali Mokhtarzadeh Conceptualization, Formal analysis, Funding acquisition, Methodology, Project administration, Resources, Supervision, Validation, Writing – review & editing, 2, *
Saeid Ghorbian Conceptualization, Formal analysis, Supervision, Validation, Writing – review & editing, 3, *
Behzad Baradaran Conceptualization, Formal analysis, Resources, Supervision, Validation, Writing – review & editing, 2
Changiz Ahmadizadeh Conceptualization, Formal analysis, Supervision, Validation, Writing – review & editing, 1
Author information:
1Department of Biology, Ah.C., Islamic Azad University, Ahar, Iran
2Immunology Research Center, Tabriz University of Medical Sciences, Tabriz, Iran
3Department of Biology, Ta.C., Islamic Azad University, Tabriz, Iran
Abstract
Introduction:
Chemotherapy drugs serve as one of the primary treatments for gastric cancer. However, challenges, including drug resistance and adverse side effects of the chemotherapy drugs like cisplatin, limit their efficacy. Long non-coding RNAs (lncRNAs) have been increasingly recognized as important regulators in oncogenesis and drug response. In this study, the potential effect of the long non-coding RNA LINC00162 (PICSAR) on modulating the chemosensitivity of gastric cancer cells to cisplatin was investigated. Furthermore, the impact of the LINC00162 silencing on cellular responses was evaluated under two conditions: after the siRNA-mediated silencing of the LINC00162 alone, and after combined treatment with cisplatin.
Methods:
Firstly, the viability of the cells following siRNA-mediated LINC00162 silencing, treatment with cisplatin, and the combination of both was evaluated. Half inhibitory concentration (IC50) of the cisplatin chemotherapy drug was assessed individually and after silencing of the LINC00162 via siRNA. Additionally, we investigated apoptosis, cell cycle arrest, migration, and colony formation in the cancer cells following the combination treatment. Finally, the effect of the combination treatment on the expression of the genes involved in these pathways, including BAX, BCL2, TP53, MMP-9, CASP3, CASP9, AKT, PI3K, and NANOG was evaluated by qRT-PCR.
Results:
Findings indicated that LINC00162 silencing increased the sensitivity of the AGS gastric cancer cells to cisplatin and reduced the IC50 of cisplatin from 19.24 µg/mL to 14.08 µg/mL. Furthermore, LINC00162 siRNA induced apoptosis, increasing the apoptosis rate to 10.69% compared to 6.19% in the control group, and also caused sub-G1 cell cycle arrest (3.94%). This effect was significantly enhanced in the combination treatment group (apoptosis: 41.7% and Sub-G1 arrest: 17.3%) compared to the control group or either single treatment. Moreover, our study demonstrated that siRNA-mediated inhibition of the LINC00162, both individually and in combination with cisplatin, decreased the migration and colony formation ability of AGS cancer cells.
Conclusion:
These findings suggest that targeting LINC00162 may potentially enhance the efficacy of cisplatin in gastric cancer cells and may represent a promising therapeutic strategy for gastric cancer.
Graphical Abstract
Keywords: LncRNA, Combination therapy, Gastric cancer, Cisplatin, Apoptosis, Metastasis
Copyright and License Information
© 2026 The Author(s).
This work is published by BioImpacts as an open access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (
http://creativecommons.org/licenses/by-nc/4.0/). Non-commercial uses of the work are permitted, provided the original work is properly cited.
Funding Statement
The authors are thankful for the support of the Immunology Research Center, Tabriz University of Medical Sciences (Grant number: 74992).
Introduction
Gastric cancer (GC), a common cancer of the gastrointestinal tract, is one of the most prevalent malignancies around the world. The 2020 GLOBOCAN report outlined GC as the 5th most prevalent cancer (with over one million new cases in 2020) and the 4th leading cause of cancer (769000 deaths in 2020) globally. Eastern Asia (Japan and Mongolia) and Eastern Europe have the highest incidence of GC.1 In recent years, remarkable advances in diagnosing and treating early GC have occurred. Although these advances have greatly improved the treatment of GC, more than half of the patients are still diagnosed at an advanced stage, with a poor prognosis.2 Diagnosis at advanced stages restricts the available treatment modalities. Surgery, chemotherapy, endoscopy, and novel targeted therapies are options for GC treatment.3,4 First-line treatments for GC are fluoropyrimidine and platinum-based chemotherapy.5
Cisplatin (cis-diamminedichloroplatinum II) is one of the common first-line chemotherapy drugs for various cancers, including GC.6 As a platinum-based chemotherapy drug, cisplatin induces cytotoxicity by forming adducts with nuclear DNA.7 Also, cisplatin increases intracellular reactive oxygen species (ROS) and this generated oxidative stress damages cancer cells. However, cancer cells can ensure their survival by maintaining a balance between oxidative stress and antioxidant activity through metabolic adaptations and modulation of their antioxidant defense mechanisms.8 In addition, various factors, including epigenetic modification, signal pathway alteration, and cell metabolism disturbance, can lead to cisplatin resistance in patients.9 Due to the limitations of platinum-based chemotherapy drugs, an unfavorable prognosis, and a high incidence of GC, ongoing investigations aim to identify novel agents for integration with standard platinum-based first-line chemotherapy to improve patient survival.5 This combination of cisplatin with other therapies may also enhance efficacy and reduce the required dose of chemotherapy drugs.
Noncoding RNAs (ncRNAs) are classified into two categories: small ncRNAs (sncRNAs) and long noncoding RNAs.10 LncRNAs are RNAs longer than 200 nucleotides that do not possess protein-coding potential.11 LncRNAs play regulatory roles at various levels, including epigenetic, transcriptional, and post-transcriptional. Several studies have reported the role of lncRNAs in cancer. Their role in cancer is categorized into four actions: signal, decoy, guide, and scaffold.12 lncRNAs can exert an oncogenic or tumor suppressor role.13-15 In gastric cancer, lncRNAs can promote cancer progression and contribute to chemoresistance. They may also serve as diagnostic and prognostic biomarkers. For instance, Luo et al reported that lncRNA EIF3J-DT induces chemoresistance in gastric cancer.16 Another study also reported the potential role of PCAT1, PCAT2, and PCAT5 in GC progression. These lncRNAs can be utilized as poor diagnostic biomarkers for GC.17 LINC00162 lncRNA, also known as PICSAR, is a novel lncRNA which was found to have aberrant expression in various malignancies, including bladder cancer, thyroid cancer, and cutaneous squamous cell carcinoma.18-21 This lncRNA exerts an oncogenic role in these malignancies and promotes cancer progression. In a study on cutaneous squamous cell carcinoma, the high expression of the PICSAR lncRNA was observed in cisplatin-resistant cells. Silencing of this lncRNA inhibited migration, invasion, and viability of the cancerous cells. It was reported that PICSAR lncRNA increased resistance through the miR-485-5p/REV3L axis.22
In this regard, we investigated the role of LINC00162 in the chemosensitivity of the GC cells to cisplatin. Furthermore, the role of LINC00162 silencing combined with cisplatin in viability, migration, colony formation, and expression of the genes involved in these biological functions was investigated. Our results indicated that siRNA-mediated silencing of the LINC00162 increased the sensitivity of the GC cells to cisplatin. Additionally, combination therapy induced apoptosis, arrested the cell cycle, and inhibited the migration of the GC cells.
Materials and Methods
Cell culture
The GC cell line (AGS) was purchased from the National Cell Bank of Iran (Pasteur Institute, Tehran, Iran) and cultured in RPMI medium (Gibco, USA) enriched with FBS (Gibco, USA) and supplemented with penicillin/streptomycin (100 U/mL and 100µg/mL, respectively). AGS cells were maintained in 25 cm2 culture flasks and were kept in an incubator (37 °C and 5% CO₂). Once the cells achieved approximately 70% confluency, they were detached using 0.25% Gibco Trypsin‐EDTA and subsequently passaged for further culture.
Transfection efficiency
AGS cells were seeded in a 6-well plate (2.5 × 105). Subsequently, the cells underwent transfection with FITC-labeled siRNA using Lipofectamine 3000 (Thermo Fisher Scientific). Efficiency and transfection success were quantified by flow cytometry (Analyser 10, Miltenyi Biotec, Germany).
Dose and time optimization of the siRNA
AGS cells were seeded at a density of 2.5 × 105 in a six-well plate and incubated for 24 h. Cells were then transfected with various doses of small-interfering RNA against lncRNA LINC00162 (synthesized by BIONEER company, Table 1), and negative control siRNA (NC, siRNA scramble without any target, sequence: 5′-UUCUCCGAACGUGUCACGUUU-3′) using Lipofectamine 3000 according to the manufacturer’s protocol. LINC00162 expression levels were measured using quantitative reverse transcription polymerase chain reaction (qRT-PCR) at 24, 48, and 72 hours post-transfection to identify the optimal time for the following assays.
Table 1.
siRNA LINC00162 sequence
|
lncRNA
|
Strand
|
Sequences
|
| LIINC00162 |
Sense |
CUCAGACAUCUGCAGUCACUUCACA |
|
|
Antisense |
UGUGAAGUGACUGCAGAUGUCUGAGGA |
RNA extraction and qRT -PCR
Total RNA was extracted using the Trizol kit (GeneAll, Korea), and RNA purity was assessed by measuring absorbance at 260/280 nm with a NanoDrop spectrophotometer (Thermo Fisher Scientific, USA). Complementary DNA (cDNA) was synthesized using the AddScript cDNA Synthesis Kit in a Bio-Rad thermal cycler (USA). Relative mRNA expression of the genes listed in Table 2 was quantified by qRT-PCR (Roche, Switzerland) with GAPDH as the internal control. The primer sequences were verified for specificity using NCBI’s Primer-BLAST tool.
Table 2.
Primer sequences
|
Primers
|
Sequences
|
|
BAX
|
Forward: 5′ GACTCCCCCCGAGAGGTCTT 3′
Reverse: 5′ ACAGGGCCTTGAGCACCAGTT 3′ |
|
BCL2
|
Forward: 5′ CTGTGGTCCACCTGACCCTCCGC 3′
Reverse: 5′ CGTACAGTTCCACAAAGGCATCCCAGC 3 |
|
CASP3
|
Forward: 5′ GGAAGCGAATCAATGGACTCTGG 3′
Reverse: 5′ GCATCGACATCTGTACCAGACC 3 |
|
CASP9
|
Forward: 5′ CCAGAGATTCGCAAACCAGAGG 3′
Reverse: 5′ GAGCACCGACATCACCAAATCC 3′ |
|
MMP-9
|
Forward: 5′ CTTTGACAGCGACAAGAAGT 3′
Reverse: 5′ AGTGAAGCGGTACATAGGGT 3′ |
|
AKT
|
Forward: 5′ AGAACGACCAAAGCCAAACACA 3′
Reverse: 5′ AGTCTGTCTGCTACAGCCTGG 3′ |
|
PI3K
|
Forward: 5′ GAAGCACCTGAATAGGCAAGTCG 3′
Reverse: 5′ GAGCATCCATGAAATCTGGTCGC 3′ |
|
TP53
|
Forward: 5′ CCTCAGCATCTTATCCGAGTGG 3′
Reverse: 5′ TGGATGGTGGTACAGTCAGAGC 3′ |
|
NANOG
|
Forward: 5′ CTAAGAGGTGGCAGAAAAACA 3′
Reverse: 5′ CTGGTGGTAGGAAGAGTAAAGG 3′ |
|
LINC00162
|
Forward: 5′ GCTCTAACTCAGGGCTCCA 3′
Reverse: 5′ TGCTCCCCACCTAAGCAATG 3′ |
|
GAPDH
|
Forward: 5′ CAAGATCATCAGCAATGCCT 3′
Reverse: 5′ GCCATCACGCCACAGTTTCC 3′ |
MTT assay
To determine the half inhibitory concentration (IC50) of cisplatin (Sigma-Aldrich, Germany) and the effects of the lncRNA LINC00162 silencing on the viability and sensitivity of AGS cells to cisplatin, an MTT assay was conducted. For this purpose, the cells were seeded in a 96-well plate (10 × 103 cells per well) and incubated for 24 h, then treated with cisplatin (3–50 µg/mL; Sigma-Aldrich, Germany) for another 24 h. Then, the complete medium and MTT solution (100 µL of complete medium and 50 µL of 2 mg/mL MTT, Sigma-Aldrich, Germany) were added to each well, and the plate was incubated for 4 hours. Formazan crystals were dissolved in 100 µL DMSO, followed by a 30 min incubation period. The absorbance of each well was determined at 570 nm with background subtraction at 620 nm, using an ELISA reader (Tecan, Switzerland). Additionally, MTT assays were also performed to measure the viability of the cells following LINC00162 siRNA transfection alone and in combination with cisplatin.
Annexin/PI apoptosis assays
Apoptosis was evaluated using the Annexin V/PI staining kit (Immunostep, Spain) in four groups: (1) LINC00162 siRNA, (2) cisplatin, (3) siRNA + cisplatin, and (4) control (untreated and untransfected). AGS cells were seeded in a 6-well plate (2.5 × 105/well). Twenty-four hours after incubation, LINC00162 siRNA was transfected into AGS cells based on the optimal concentration and incubation time, following the manufacturer's protocol for Lipofectamine 3000. Cisplatin was applied to the drug and combination groups the next day, and the plate was incubated for 24 h. Subsequently, the cells were detached with trypsin/EDTA and stained with Annexin V/PI. Apoptosis rate in each group was evaluated, and the data were analyzed using FlowJo.
DAPI staining
Apoptosis was further examined by DAPI (4′,6‐diamidino‐2‐phenylindole) staining to visualize characteristic morphological changes such as chromatin condensation and nuclear fragmentation. Briefly, AGS cells (10 × 103/well) were seeded in a 96-well plate, transfected with LINC00162 siRNA, and treated with cisplatin after 24 h. Twenty-four hours after treatment, the cells of each group were fixed with 4% paraformaldehyde. Cells were washed multiple times with PBS and permeabilized with 0.1% Triton X-100 for 15 minutes. Another PBS wash was performed in the following step, and fixed cells were stained with 100 µL of 0.1% DAPI solution (Sigma-Aldrich, MO). Morphologic changes of each group were monitored by the Cytation 5 fluorescence imaging system (BioTK).
Cell cycle assay
Cell cycle profile of the AGS cells following LINC00162 silencing and cisplatin treatment was assessed by flow cytometry. A total of 2.5 × 105 cells were seeded into each well of a 6-well plate. After 24 hours, LINC00162 siRNA was transfected into the related groups, and after 24 h, the cells were treated with cisplatin. Cells were collected after 24 h of incubation and subsequently fixed overnight in 80% ethanol at −20 °C. The cells were incubated for 30 minutes in 500 µL of PBS containing 1 mg/mL RNase A. Afterward, the pellet was resuspended in 500 µL of cold PBS containing DAPI and Triton. Cell cycle arrest and progression were examined using flow cytometry, and data were analyzed by FlowJo.
Wound healing assay
The wound healing assay, also known as the scratch assay, was used to determine the migration of the AGS cancer cells. For this purpose, AGS cells (1.5 × 105/well) were seeded in six-well plates, transfected with LINC00162 siRNA, and treated with cisplatin. A wound in the culture cells was simulated by creating a scratch across the surface of the cell monolayer. A sterile pipette tip created this scratch. The plates were subsequently imaged using an inverted light microscope at 0, 24, and 48 h after treatment. Cell migration was assessed by observing the accumulation of cells within the gap area and comparing treatment groups to the control group.
Colony formation assay
The ability of AGS cells to form colonies after the siRNA-mediated silencing of LINC00162 lncRNA and treatment with cisplatin, compared with the control group, was assessed using a colony formation assay. For this assay, we used a 12-well plate (3 × 10³ cells per well), which was incubated for 10 days following transfection with siRNA and treatment with cisplatin. After an incubation period, the cells were washed with PBS and fixation of the AGS cells was performed using 4% paraformaldehyde. Fixed cells were stained with 0.5% crystal violet for 30 minutes. Following a final wash, formed colonies in each experimental group were photographed.
Statistical analysis
Statistical analysis was performed using GraphPad Prism software (Version 9, CA, USA). The experiments were conducted in technical triplicate, and all data are presented as mean ± standard error (SE). T-test and one-way ANOVA were used to compare the means between experimental groups. Statistical significance was considered as a p-value less than 0.05.
Results
siRNA was successfully transfected into AGS cells
Quantitative assessment via flow cytometry demonstrated that FITC-labelled siRNA was successfully internalized by the AGS cell line. The measured transfection efficiency reached 88.1%, a significant increase over the untransfected control. These data confirm the high efficacy of Lipofectamine and the successful transfection of the FITC-labelled siRNA into AGS cells (Fig. 1A).
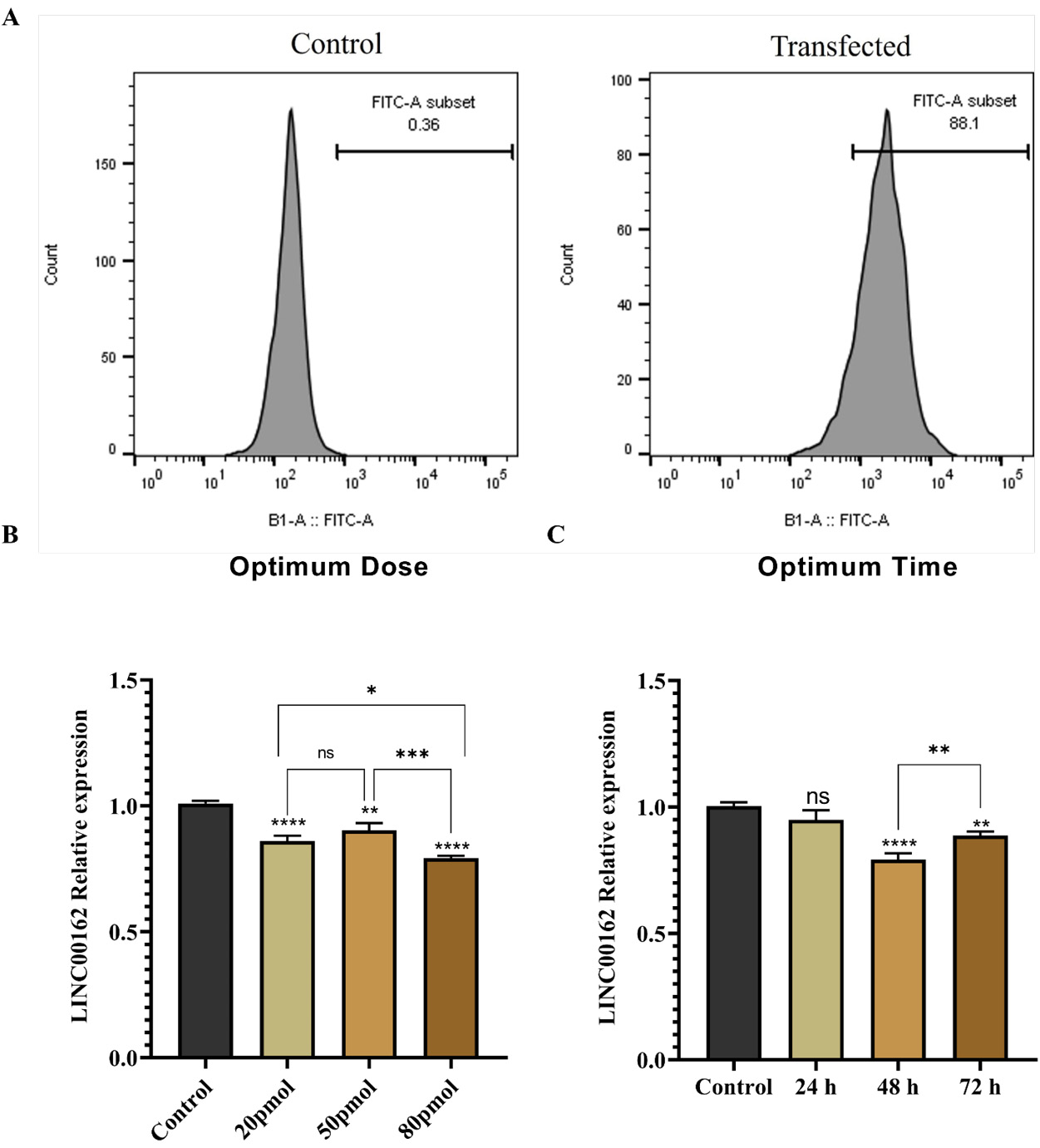
Fig. 1.
A. Successful introduction of the FITC-labeled siRNA into AGS cells was confirmed with high delivery rate of 88.1%. B. The most notable decrease in LINC00162 expression within the AGS cell line was observed at 80 pmol concentration. C. siRNA transfection with 80 pmol suppressed LINC00162 expression effectively 48 h after transfection (****P < 0.0001, ***P < 0.001, **P < 0.01, *P < 0.05, and ns = not significant).
.
A. Successful introduction of the FITC-labeled siRNA into AGS cells was confirmed with high delivery rate of 88.1%. B. The most notable decrease in LINC00162 expression within the AGS cell line was observed at 80 pmol concentration. C. siRNA transfection with 80 pmol suppressed LINC00162 expression effectively 48 h after transfection (****P < 0.0001, ***P < 0.001, **P < 0.01, *P < 0.05, and ns = not significant).
LINC00162 siRNA transfection reduced LINC00162 expression in AGS cells
To determine the optimal dose for the transfection, AGS cells were transfected with varying doses of LINC00162 siRNA (20, 50, and 80 pmol). According to qRT-PCR results (Fig. 1B), a significant reduction in LINC00162 expression was seen in all doses. While there was no significant difference between 20 and 50 pmol, 80 pmol reduced the expression of LINC00162 more significantly compared to the untransfected control group (95% CI = 0.166 to 0.267, P < 0.0001), 20 pmol (95% CI = 0.0163 to 0.117, P < 0.05), and 50 pmol (95% CI = 0.05932 to 0.1607, P < 0.001). Also, among various transfection times, including 24, 48, and 72 h, 48 h was confirmed to be the best period for siRNA activity (Fig. 1C). 80 pmol and 48 h were determined as the effective dose and time for the subsequent experiments.
siRNA-mediated suppression of LINC00162 reduced the viability of AGS cells
MTT assay showed the viability of the AGS cells following each treatment. The data obtained indicated that transfection with NC had no significant effect on cell viability compared to the untransfected control group. Silencing of the LINC00162 via siRNA significantly reduced AGS cell viability (Fig. 2A, 95% CI = 11.06 to 32.57, P < 0.01 and 95% CI = 3.113 to 24.62, P < 0.05, respectively) compared to the control and NC group.
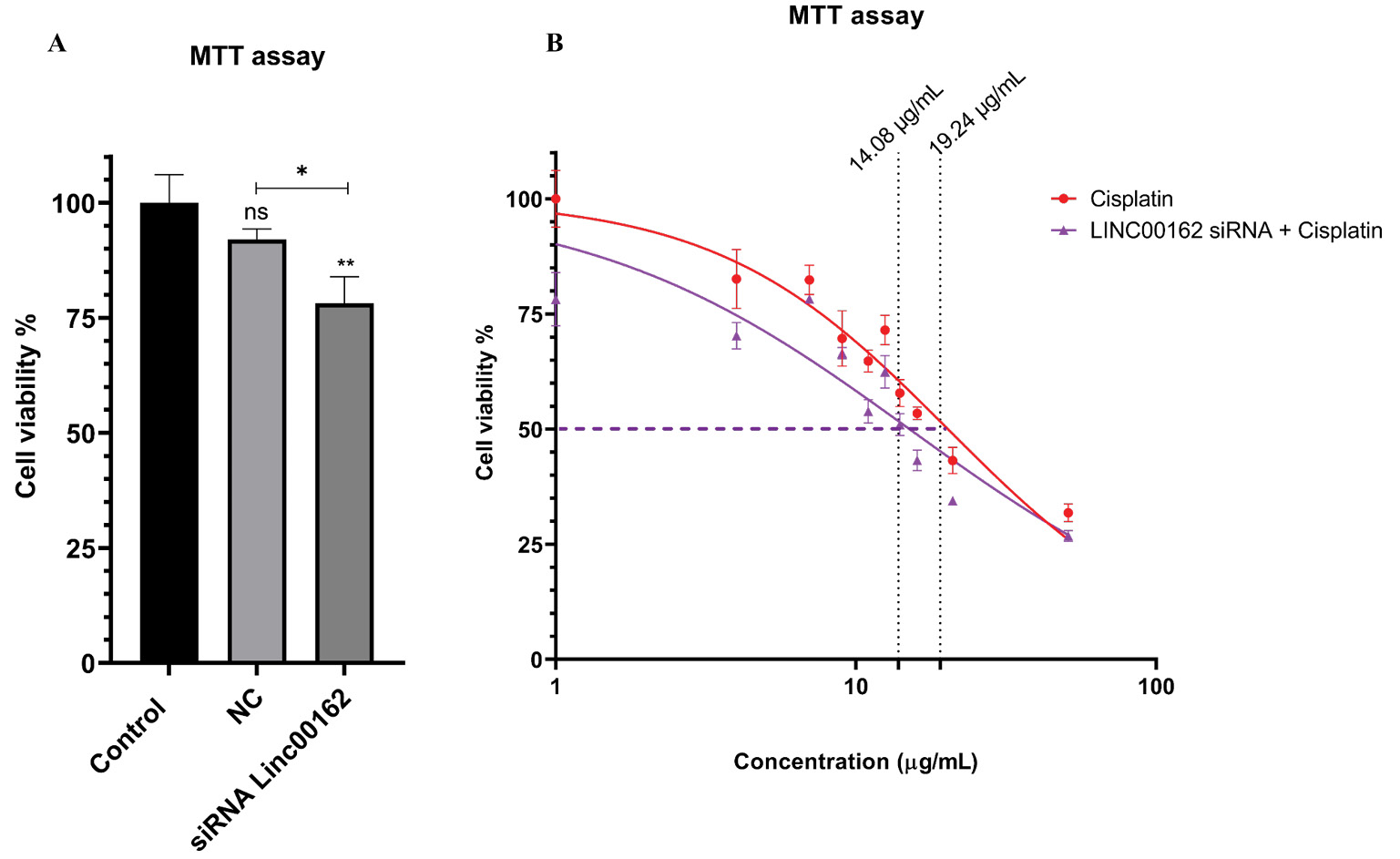
Fig. 2.
MTT assay evaluated the viability of the cells after each treatment. (A) Suppression of LINC00162 expression significantly reduced the viability of the AGS cells (**P < 0.01, *P < 0.05, and ns = not significant).(B) The MTT assay revealed that siRNA-mediated LINC00162 silencing increased the sensitivity of the AGS cells to the cisplatin chemotherapeutic drug and decreased the IC50 of the drug from 19.24 µg/mL to 14.08 µg/mL.
.
MTT assay evaluated the viability of the cells after each treatment. (A) Suppression of LINC00162 expression significantly reduced the viability of the AGS cells (**P < 0.01, *P < 0.05, and ns = not significant).(B) The MTT assay revealed that siRNA-mediated LINC00162 silencing increased the sensitivity of the AGS cells to the cisplatin chemotherapeutic drug and decreased the IC50 of the drug from 19.24 µg/mL to 14.08 µg/mL.
LINC00162 lncRNA inhibition increased cisplatin sensitivity in AGS cells
The IC50 value of cisplatin was assessed using the MTT assay, both for the drug alone and after silencing LINC00162 via siRNA. For this purpose, the AGS cells were treated with various doses of cisplatin (3 to 50 µg/mL). The viability of the AGS cells decreased in a dose-dependent manner following treatment with cisplatin. While the IC50 of the cisplatin alone was reported as 19.24 µg/mL, transfection of LINC00162 siRNA reduced the IC50 of the cisplatin to 14.08 µg/mL. The results indicate that siRNA-mediated silencing of the LINC00162 increased the sensitivity of the AGS cells to cisplatin, resulting in reduced IC50 dose (Fig. 2B).
LINC00162 silencing significantly enhanced the apoptotic effects of cisplatin in AGS cells
The induction of apoptosis following LINC00162 silencing and cisplatin treatment was investigated using AnnexinV/PI staining. Following transfection with LINC00162 siRNA, AGS cells were treated with a dose less than IC50 of cisplatin. Compared to the control group, which showed a 6.19% apoptosis rate, both single treatments increased cell death. This rate increased to 10.69% following LINC00162 siRNA transfection and to 16.40% after cisplatin treatment. These findings indicate that combination of the LINC00162 silencing and cisplatin treatment enhanced apoptosis rate to 41.7% and significantly increased apoptosis induction compared to the control group (P < 0.0001) and individual treatments (P < 0.0001) (Fig. 3A).
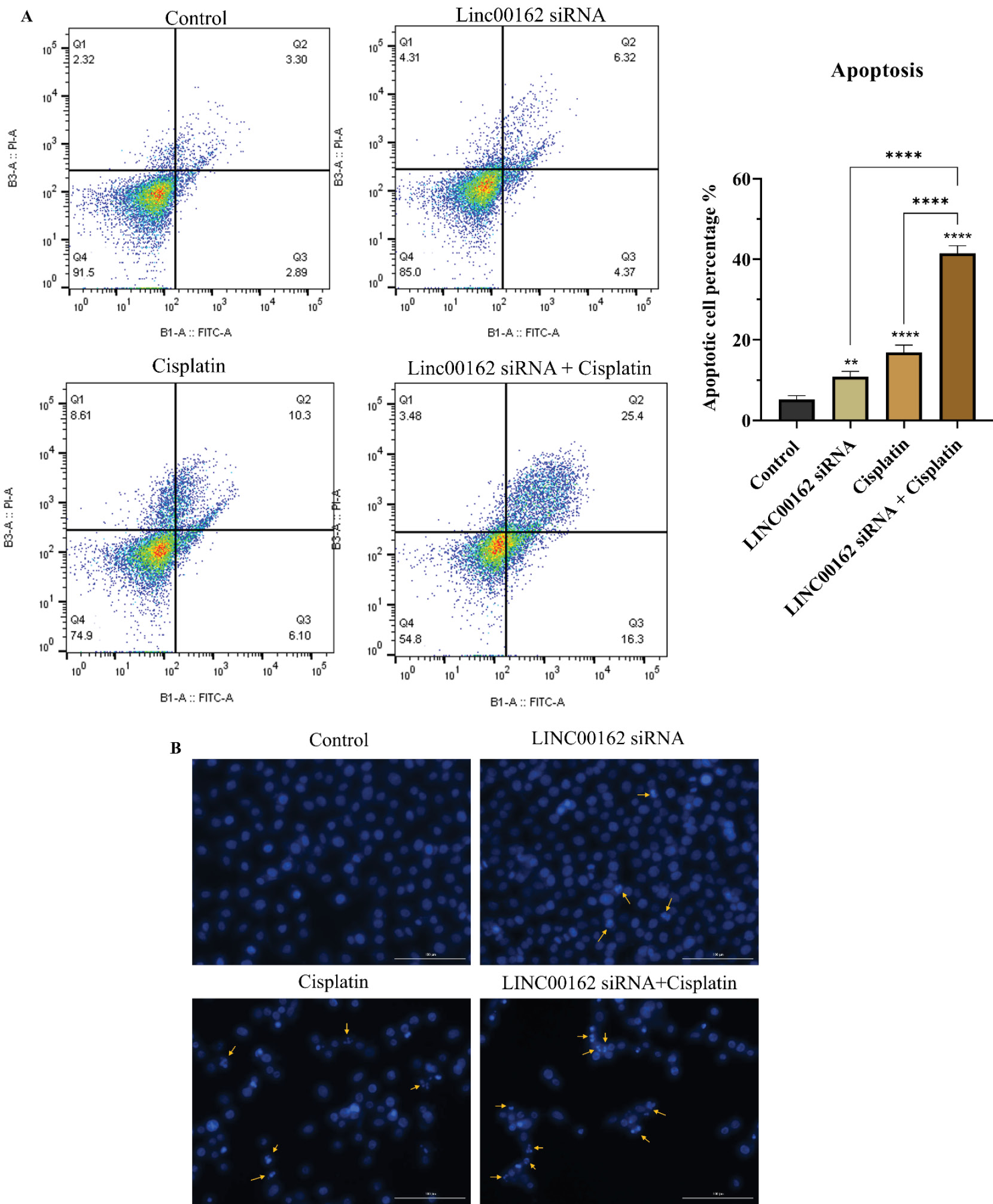
Fig. 3.
Combined treatment promotes apoptosis in AGS cells. (A) Combination of LINC00162-siRNA and cisplatin treatment significantly increased apoptosis rate in AGS cells (**P < 0.01, ****P < 0.0001, and ns = not significant). (B) DAPI staining was utilized to track chromatin fragmentation.
.
Combined treatment promotes apoptosis in AGS cells. (A) Combination of LINC00162-siRNA and cisplatin treatment significantly increased apoptosis rate in AGS cells (**P < 0.01, ****P < 0.0001, and ns = not significant). (B) DAPI staining was utilized to track chromatin fragmentation.
DAPI staining was also conducted to observe chromatin fragmentation of AGS cells following LINC00162 silencing and cisplatin treatment. The obtained results showed more fragmented cells in the group that received the combination treatment relative to those treated with either agent individually or the untreated control group (Fig. 3B).
Also, the investigation of apoptotic gene expression using the qRT-PCR assay showed a slight upregulation in the expression of the BAX following siRNA transfection; this change was not statistically significant (95% CI = -1.134 to 0.1809, ns). While cisplatin treatment significantly increased expression of the BAX (95% CI = -1.738 to -0.422, P < 0.01), a significant increase in the fold change levels of the BAX was observed following combination treatment (95% CI = -6.251 to -4.936, P < 0.0001) (Fig. 4A). Furthermore, LINC00162 silencing did not significantly alter the expression of CASP3 relative to the control group. In contrast, the same treatment significantly upregulated CASP9 (95% CI = -1.183 to -0.0972, P < 0.05). Also, cisplatin treatment significantly increased the expression of the CASP3 (95% CI = -2.189 to -0.817, P < 0.001) and CASP9 (95% CI = -1.469 to -0.383, P < 0.01) genes compared to the untreated control group. Furthermore, significant upregulation in the expression of CASP3 and CASP9 compared to individual treatments and the control group was observed following combined silencing of LINC00162 and treatment with cisplatin (Fig. 4C and 4D). Furthermore, BCL2 mRNA levels were significantly reduced in response to both LINC00162 silencing (95% CI = 0.356 to 0.4974, P < 0.0001) and cisplatin treatment (95% CI = 0.746 to 0.887, P < 0.0001). Also, the combination treatment resulted in a significantly greater decrease in Bcl-2 expression than LINC00162 silencing or cisplatin treatment alone. (Fig. 4B).
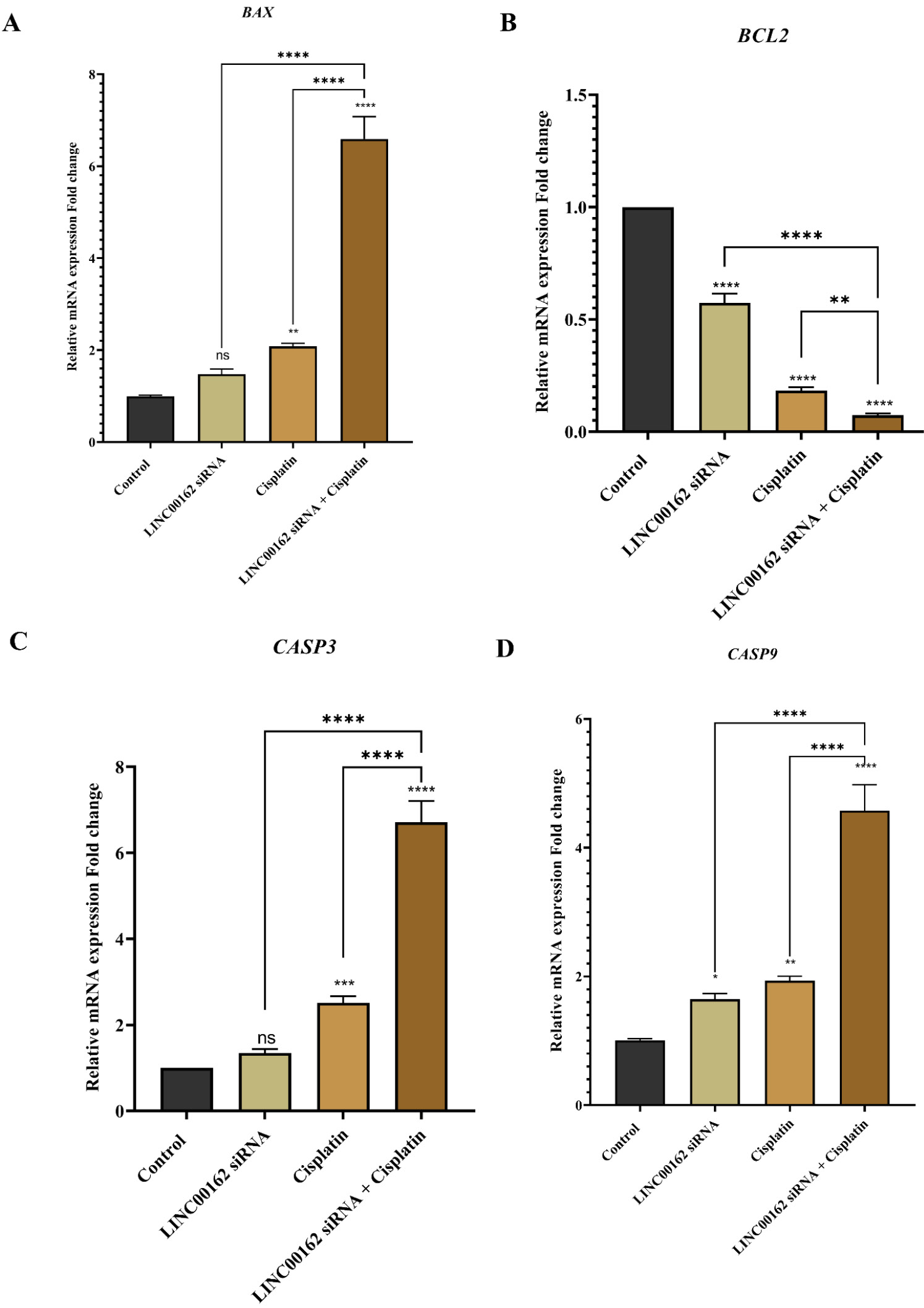
Fig. 4.
Effect of combination treatment on the expression of the apoptosis-related genes. Combination therapy significantly increased the expression of the BAX, CASP3, CASP9, and decreased Bcl-2 mRNA levels (*P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, and ns = not significant).
.
Effect of combination treatment on the expression of the apoptosis-related genes. Combination therapy significantly increased the expression of the BAX, CASP3, CASP9, and decreased Bcl-2 mRNA levels (*P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, and ns = not significant).
The combination of LINC00162 silencing and cisplatin treatment induced cell cycle arrest at the Sub-G1 phase in AGS cells
Cell cycle arrest following each treatment and their combination was investigated by Flow cytometry. According to the results, siRNA-mediated downregulation of the LINC00162 increased the population of the sub-G1 arrested cells from 1.46% in the control group to 3.94%. Cisplatin treatment increased the percentage of the cells arrested in the Sub-G1 phase to 11.3%. Additionally, the combination of both treatments significantly increased the number of cells in the sub-G1 phase, reaching 17.3%. The percentage of arrested cells in the combination treatment group was significantly higher compared to the siRNA transfection and cisplatin treatment alone (95% CI = -16.43 to -11.59, P < 0.0001 and 95% CI = -8.775 to -3.932, P < 0.001, respectively) and the control group (95% CI = -18.87 to -14.03, P < 0.0001). The results showed that cisplatin and siRNA LINC00162 combination therapy increase Sub-G1 arrested cells (Fig. 5).
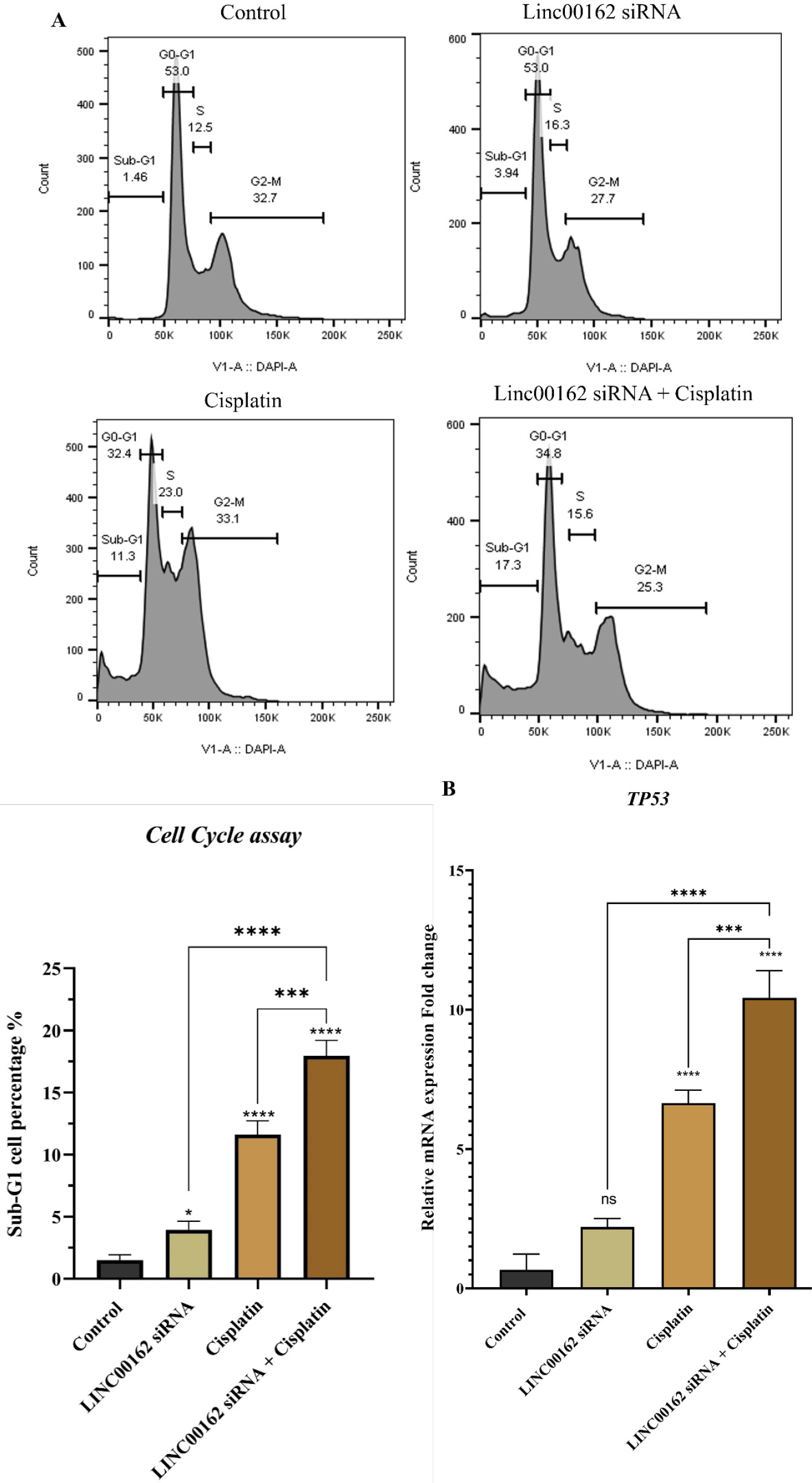
Fig. 5.
The effect of LINC00162-siRNA and cisplatin combination on the cell cycle progression. (A) siRNA-mediated downregulation of the LINC00162, combined with the cisplatin treatment, arrested AGS cells in the sub-G1 phase. (B) Combined group of LINC00162 and cisplatin significantly increased the mRNA level of the TP53 gene compared to individual treatment and the control group (*P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, and ns = not significant)
.
The effect of LINC00162-siRNA and cisplatin combination on the cell cycle progression. (A) siRNA-mediated downregulation of the LINC00162, combined with the cisplatin treatment, arrested AGS cells in the sub-G1 phase. (B) Combined group of LINC00162 and cisplatin significantly increased the mRNA level of the TP53 gene compared to individual treatment and the control group (*P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, and ns = not significant)
qRT-PCR was used to evaluate mRNA levels of the TP53 gene. As shown in Fig. 5B, although LINC00162 silencing increased the expression of the TP53 gene, this upregulation did not reach statistical significance relative to the control group (95% CI = -3.172 to 0.098). Additionally, a significant increase in the expression of the TP53 was observed in the cells treated with cisplatin (95% CI = -7.619 to -4.348, P < 0.0001) and those that received combination treatment (95% CI = -11.40 to -8.128, P < 0.0001).
LINC00162 downregulation and cisplatin treatment decrease the migration ability of the AGS cells
A wound healing assay was performed to assess the migration ability of the AGS gastric cancer cells following each treatment. A significant reduction in migration and viability of AGS cells was observed 48 hours following co-treatment with LINC00162 siRNA and cisplatin (Fig. 6A). The migration of the cells transfected with LINC00162 siRNA was also reduced compared to the control group. However, this reduction was more in the cisplatin-treated alone and the combination group (Fig. 6A).
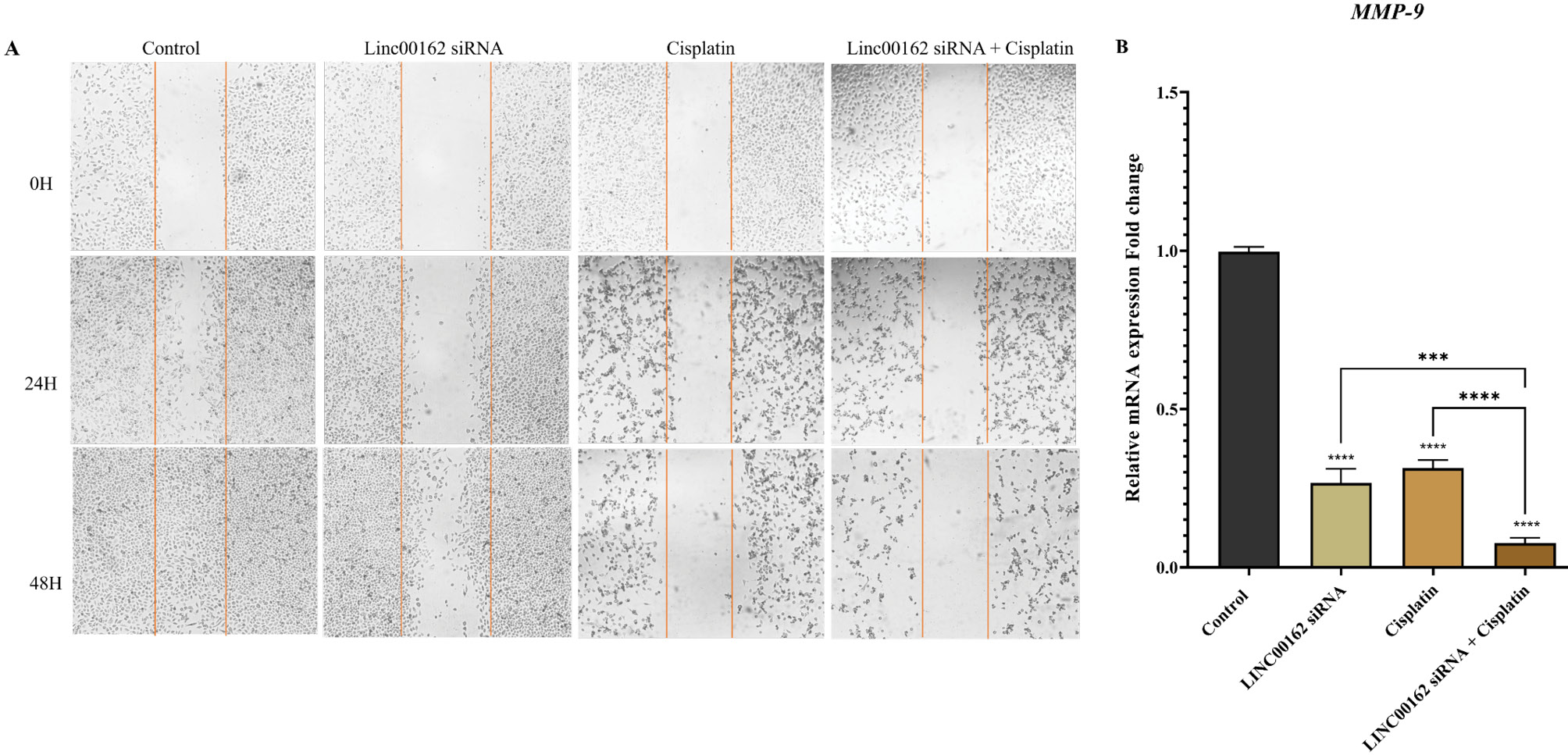
Fig. 6.
Effect of combined LINC00162 silencing and cisplatin on cell migration. (A) After 48h, siRNA-mediated silencing of the LINC00162 combined with cisplatin significantly reduced the migration of the AGS cells. (B) qRT-PCR results illustrated that LINC00162 and cisplatin cooperatively downregulated the expression levels of metastasis-related genes MMP-9 (***P < 0.001, ****P < 0.0001, and ns = not significant).
.
Effect of combined LINC00162 silencing and cisplatin on cell migration. (A) After 48h, siRNA-mediated silencing of the LINC00162 combined with cisplatin significantly reduced the migration of the AGS cells. (B) qRT-PCR results illustrated that LINC00162 and cisplatin cooperatively downregulated the expression levels of metastasis-related genes MMP-9 (***P < 0.001, ****P < 0.0001, and ns = not significant).
To confirm scratch assay findings, we assessed the expression of the MMP9 gene, a key regulator of cell migration. qRT-PCR analysis revealed that both LINC00162 silencing (95% CI = 0.656 to 0.804, P < 0.0001) and cisplatin treatment (95% CI = 0.609 to 0.757, P < 0.0001) significantly reduced MMP9 expression in AGS cells. Moreover, the combination of these therapeutic approaches led to a markedly greater suppression of MMP9 expression compared to the control group (95% CI = 0.846 to 0.993, P < 0.0001) and each treatment group (P < 0.001 and P < 0.0001) (Fig. 6B).
LINC00162 downregulation and cisplatin treatment reduced the colony formation ability of the AGS cells
The clonogenic ability of AGS cells was evaluated using a colony formation assay following treatment with LINC00162 siRNA, cisplatin, or a combination of both. Results indicated that LINC00162 transfection slightly reduced the number of colonies. Also, chemotherapy with cisplatin significantly reduced the number of colonies formed and their size. However, the number and the size of the colonies in the cells treated with both LINC00162 siRNA and cisplatin were significantly reduced compared to individual treatments (Fig. 7A).
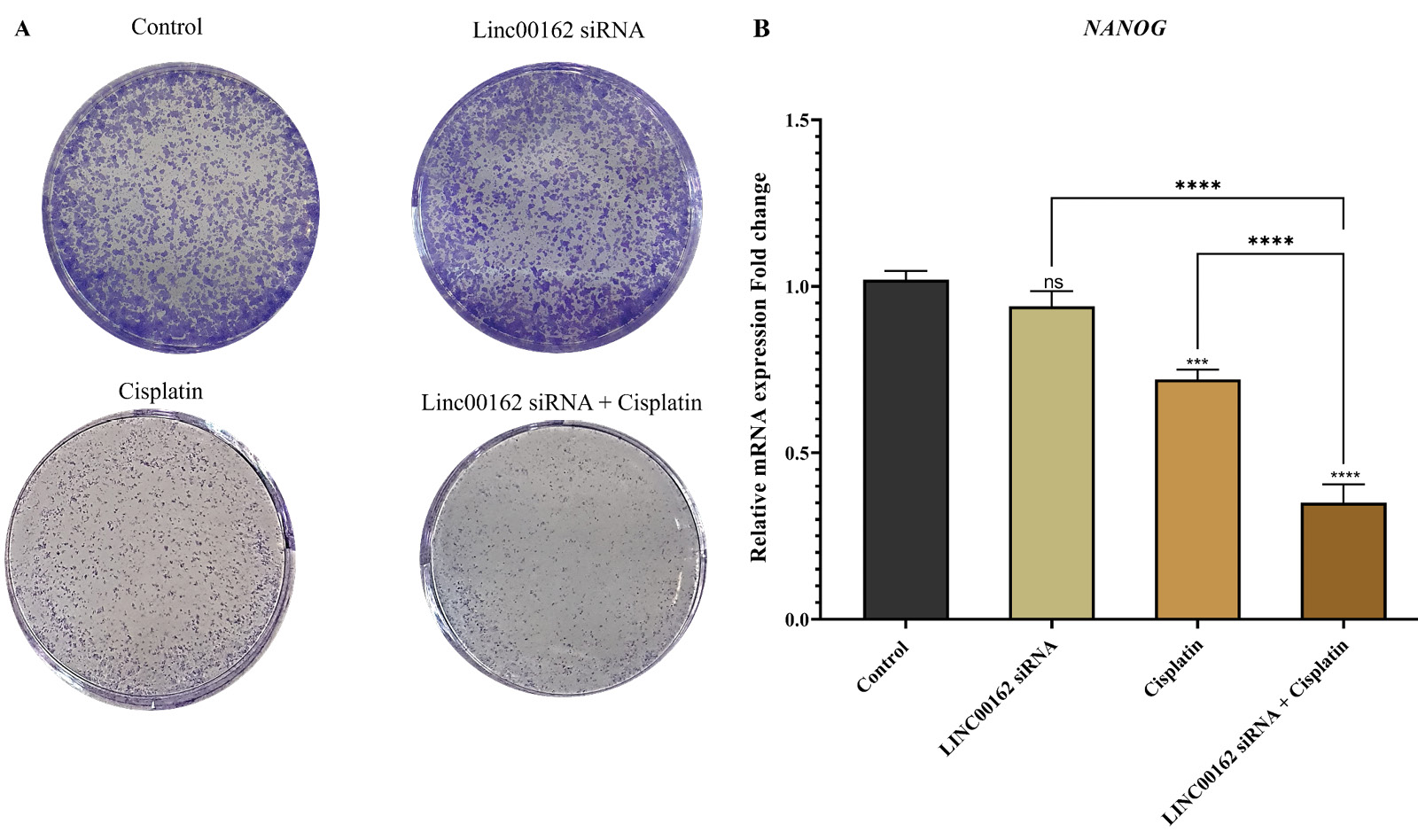
Fig. 7.
Effect of the combination therapy on the clonogenic ability of AGS cells and expression of the NANOG gene. (A) LINC00162 silencing and cisplatin treatment reduced colony number and size. (B) Effect of the siRNA-mediated silencing of the LINC00162 and cisplatin treatment on the expression of the NANOG (***P < 0.001, ****P < 0.0001, and ns = not significant).
.
Effect of the combination therapy on the clonogenic ability of AGS cells and expression of the NANOG gene. (A) LINC00162 silencing and cisplatin treatment reduced colony number and size. (B) Effect of the siRNA-mediated silencing of the LINC00162 and cisplatin treatment on the expression of the NANOG (***P < 0.001, ****P < 0.0001, and ns = not significant).
For confirmation of colony assay results, we evaluated expression of the NANOG, which is a key stem cell marker. Although LINC00162 siRNA decreased the expression of this gene, these changes were not significant compared to untreated and untransfected cells (95% CI = −0.028 to 0.188). Cisplatin treatment of the cells significantly reduced the expression of NANOG compared to the control group (95% CI = 0.192 to 0.408; P < 0.0001). AGS cells that received combination therapy, however, displayed a greater decrease in expression of this gene (95% CI = 0.562 to 0.778; P < 0.0001). The findings suggest that siRNA-mediated downregulation of LINC00162 increases the sensitivity of cells to cisplatin, which, in combination with this chemotherapy drug, decreases the stemness ability of the AGS gastric cancer cells (Fig. 7B).
Cisplatin treatment combined with LINC00162 downregulation decreased AKT and PI3K expression
We further assessed the expression level of the AKT gene, a key regulator implicated in cell survival, metastasis, and chemosensitivity in GC. Both LINC00162 (95% CI = 0.0228 to 0.264, *P < 0.05) silencing and cisplatin treatment (95% CI = 0.129 to 0.370, ***P < 0.001) independently reduced AKT mRNA expression compared to the control group (Fig. 8). Notably, the combination therapy resulted in a more significant downregulation of AKT expression than either treatment alone or the control group (95% CI = 0.499 to 0.7405, P < 0.0001, Fig. 8A). Furthermore, a modest reduction in the expression of the PI3K was observed following lncRNA silencing; however, this reduction was not statistically significant. Cisplatin treatment also significantly reduced the expression of PI3K. Additionally, when cisplatin treatment was combined with LINC00162 silencing, expression of PI3K was reduced considerably compared with either treatment alone (Fig. 8B).
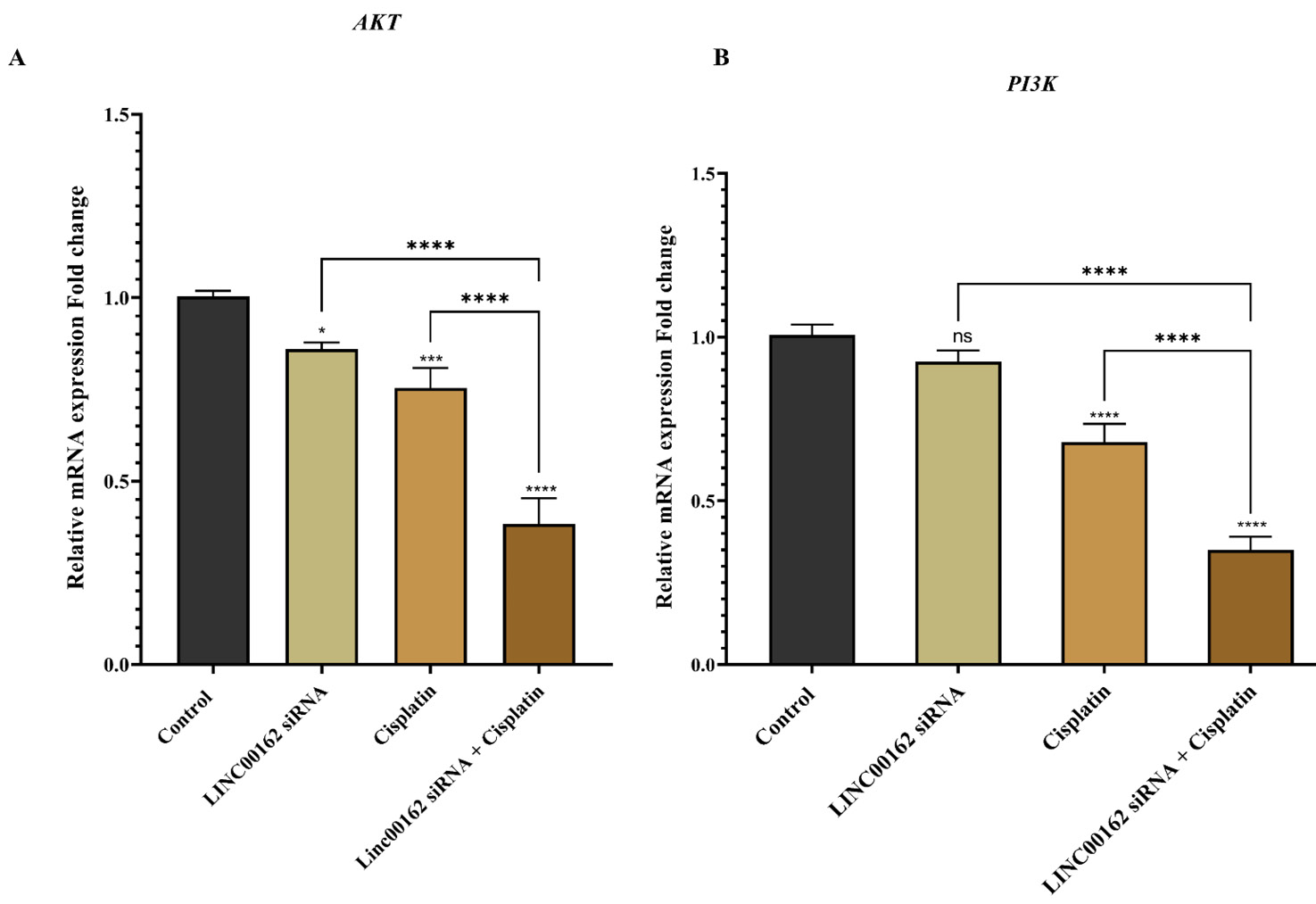
Fig. 8.
AKT and PI3K mRNA expression levels following LINC00162-siRNA transfection and cisplatin treatment. Combination therapy significantly decreased AKT and PI3K mRNA levels (*P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, and ns = not significant).
.
AKT and PI3K mRNA expression levels following LINC00162-siRNA transfection and cisplatin treatment. Combination therapy significantly decreased AKT and PI3K mRNA levels (*P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, and ns = not significant).
Discussion
Gastric cancer, with a 32.4% survival rate, is the fourth most deadly cancer all around the world.23 It is a heterogeneous and aggressive disease resulting from multiple interactions among genetic, environmental, and host factors.24 Depending on the tumor stage, treatment effectiveness and treatment strategy will differ. Metastasis can occur in advanced gastric cancers, which reduces the overall prognosis. In recent years, extensive research efforts have focused on enhancing the prognosis of GC patients, leading to the development of neoadjuvant chemotherapy, radiotherapy, and molecular-targeted therapies as effective treatment strategies.25 Cisplatin, a platinum-based chemotherapy drug, is being utilized against various cancers and is a first-line drug for patients with advanced GC.26 Cisplatin disrupts the structure of the DNA by forming interstrand and intrastrand crosslinks. Cellular proteins repair these DNA damages. Increased repair of these damages can lead to cisplatin resistance.27 A considerable proportion of cancer patients eventually develop resistance to cisplatin. Intrinsic and extrinsic drug resistance can reduce the effectiveness of chemotherapy drugs. Mechanisms of chemoresistance include increased drug efflux, the ability of the GC cells to repair anticancer drug-induced DNA damage, and modulation of apoptotic genes.28 Hence, investigating underlying mechanisms and developing novel strategies is essential to overcome chemoresistance.
Various studies indicated that lncRNAs play a key role in tumor physiology and pathology by regulating the expression of various genes and modulating biological pathways, including apoptosis, cell cycle, migration, and metastasis.29 Studies also investigated the role of the lncRNAs in the chemosensitivity of the cancerous cells. For instance, a recent study conducted in 2025 reported increased sensitivity to 5-FU following silencing of the LINC02323 in gastric cancer cells. This lncRNA negatively regulates miR-139-3p and, in this way, modulates the sensitivity of the gastric cancer cells to 5-FU.30 LncRNA DLEU2, which is upregulated in gastric cancer cells and tumors, is involved in Taxol resistance of gastric cancer cells. Bioinformatic analysis indicated that this lncRNA sponges miR-30c-5p. Downregulation of the LncRNA DLEU2 or overexpression of the miR-30c-5p increased Taxol sensitivity of gastric cancer cells.31 Wang et al reported that downregulation of ROR lncRNA in gastric cancer cells reduced expression of the MRP1 and MDR genes and increased apoptosis. Also, depletion of ROR lncRNA increased the sensitivity of the cells to Adriamycin and Vincristine.32 Furthermore, another study reported that upregulation of HOTAIR lncRNA promotes cisplatin resistance in gastric cancer cells, while reducing the expression of HOTAIR, sensitizes gastric cancer cells to cisplatin.33 In our study conducted on thyroid cancer cells, LINC00162 silencing increased the sensitivity of cancerous cells to sorafenib and increased the apoptosis rate in cancer cells individually and combined with chemotherapy drugs.34 Wang et al demonstrated the upregulation of LINC00162 in cutaneous squamous cell carcinoma cells. Based on their findings, LINC00162 knockdown suppressed cisplatin resistance in cisplatin-resistant cells and in vivo. Their study demonstrated that reducing LINC00162 expression could diminish resistance to cisplatin in these cancer cells.22 Previously described study by Wang et al on the effect of LINC00162 in cutaneous squamous cell carcinoma cells and its impact on the cisplatin resistance of the cancer cells, paved the way for us to study this lncRNA further. Our study is the first to demonstrate the functional role of LINC00162 in modulating chemosensitivity to cisplatin in gastric cancer, thereby extending the understanding of this lncRNA across different cancer types and tissue origins. Furthermore, we provide a comprehensive mechanistic investigation by examining multiple cellular processes, including apoptosis induction, cell cycle arrest, migration inhibition, and colony formation suppression, and their associated molecular pathways involving BAX, BCL2, TP53, MMP-9, CASP3, CASP9, AKT, PI3K, and NANOG, which were not explored in previous studies.
The silencing of LINC00162 via siRNA reduced the viability of AGS gastric cancer cells. The IC₅₀ of cisplatin decreased from 19.24 µg/mL to 14.08 µg/mL following LINC00162 silencing, indicating enhanced chemosensitivity and a lower drug dose required for half-maximal inhibition. Our findings aligned with the previous study mentioned earlier in which PICSAR contributed to cisplatin resistance in cutaneous squamous cell carcinoma cells.22
We next evaluated apoptosis in AGS cells treated with LINC00162 siRNA, cisplatin, or their combination. The apoptosis rate was 6.19% in control cells, 10.69% after LINC00162 silencing, 16.40% with cisplatin, and markedly increased to 41.7% in the combined treatment group. To confirm the Annexin V/PI assay results, we analyzed apoptosis-related gene expression. One of the essential apoptosis regulators is the B-cell lymphoma 2 (BCL-2) protein family. Members of this family divide into various subgroups and play a key role in regulating tumorigenesis, cell death, and responses to anticancer therapy. This group of proteins includes the anti-apoptotic and pro-apoptotic proteins represented by BCL-2 and BAX, respectively.35 Caspase-3, a cysteine-aspartic acid protease (caspase) family member, plays a critical role in the proteolytic degradation that occurs during apoptosis. Procaspase-3, the initial form of caspase-3, can be activated by Caspase-8, Caspase-9, or Caspase-10. Studies indicated that Caspase-3 may act as a tumor suppressor in human gastric cancer.36 qRT-PCR results supported flow cytometry findings. Based on the results, LINC00162 silencing alone did not significantly alter BAX expression but significantly downregulated BCL2. Combined treatment led to a notable upregulation of BAX and downregulation of BCL2, confirming enhanced apoptosis induction. Evaluation of CASP3 and CASP9 gene expression also showed that co-treatment with LINC00162 siRNA and cisplatin significantly increased expression of these pro-apoptotic genes relative to the control and individual treatment groups. Consistent with our findings, a study conducted on Hep3B cells derived from hepatocellular carcinoma showed that knockdown of PICSAR induced apoptosis, whereas its overexpression decreased apoptosis, inhibited BAX, and promoted Bcl-2 expression.37 Additionally, silencing of LINC0162 in HeLa cells increased the number of early and late apoptotic cells.38
In addition, our study demonstrated that silencing LINC00162 in combination with cisplatin treatment significantly reduced the colony formation ability of AGS gastric cancer cells. Consistent with our findings, PICSAR knockdown inhibited colony formation of hepatocellular carcinoma and HeLa cells.37,38 The Nanog homeobox (NANOG) has attracted increasing attention as a stemness factor involved in tumorigenesis and malignancy. Based on accumulating evidence, NANOG is involved in chemoresistance, tumorigenesis, and epithelial–mesenchymal transition (EMT).39 In a study conducted in 2010, overexpression of NANOG/NANOGP8 was detected in primary gastric cancer tumors.40 Another study investigated the expression of NANOG in 100 gastric cancer tumor tissues compared to the paired adjacent normal tissues. Researchers reported overexpression of NANOG in gastric cancer tumor tissues.41 Vasefifar et al targeted Nanog expression in MKN-45 cells. They reported that MDR-1 mRNA expression was significantly reduced after Nanog downregulation, and cisplatin sensitivity was increased in MKN-45 cells.42 In our findings, while silencing of LINC00162 alone did not significantly alter the expression of the NANOG, we observed significant downregulation of the NANOG following combination of LINC00162 silencing and treatment with cisplatin.
We also analyzed cell cycle progression after individual and combined treatments. Flow cytometry revealed that LINC00162 silencing alone increased the sub-G1 cell population, while cisplatin treatment, alone or combined with LINC00162 silencing, further enhanced sub-G1 arrest. These results are consistent with the apoptosis assay, confirming that LINC00162 silencing promotes cell death in AGS cells. Wang et al reported blockade of the G0/G1 phase following LINC00162 silencing in bladder cancer cells.19 To validate our cell cycle findings, we examined TP53 expression. Tumor suppressor p53 is mutated in the early stages of gastric cancer.43 We examined that while LINC00162 silencing and cisplatin treatments individually increased expression of the TP53, this alteration was not significant in the LINC00162 siRNA-treated group. However, combination treatment significantly restored TP53 expression.
We also assessed the effects of LINC00162 silencing and cisplatin on AGS cell migration and invasion. Wound healing assays showed that both LINC00162 siRNA and cisplatin individually reduced cell migration, with the combination treatment producing a more pronounced inhibitory effect. Matrix metalloproteinases (MMPs), which break down proteins in the structure of the extracellular matrix (ECM), are involved in various processes, including cell migration.44 Liu et al reported the accumulation of tumor-associated macrophages (TAMs) in gastric cancer. They reported that TAMs promote gastric cancer metastasis by secreting MMP-9, which induces metastasis via the PI3K/AKT/Snail signaling pathway. Inhibition of MMP-9 decreased metastasis through suppression of the EMT process.45 In this regard, we evaluated the expression of the MMP-9 gene. Although both lncRNA silencing and cisplatin treatment decreased mRNA levels of the MMP-9 gene, this downregulation was significantly more in the combination treatment group. Similar to our findings, a study conducted in cutaneous squamous cell carcinoma cell lines reported a significant decrease in cell migration following knockdown of PICSAR expression.46 Also, Luo et al's study in gastric cancer cells showed downregulation of the MMP-9 and reduced progression of the gastric cancer cells following knockdown of the LINC00483.47
The PI3K/Akt/mTOR pathway is one of the essential pathways playing a role in metastasis, survival, and resistance to chemotherapy in gastric carcinoma.48 As mentioned earlier, MMP-9 can induce metastasis of gastric cancer cells through the PI3K/AKT/Snail signaling pathway. The role of AKT in the chemosensitivity of gastric cancer cells to cisplatin was also demonstrated by Tao et al. Their findings indicated that inhibition of the AKT using MK-2206 enhances the cytotoxic effect of cisplatin and increases the apoptosis rate in AGS cells.49 Furthermore, a study reported higher expression of PI3K/AKT/mTOR signaling pathway proteins in cisplatin-resistant AGS gastric cancer cells. Silencing of the LncRNA CCAT1 increased the sensitivity of the AGS cells to cisplatin, induced apoptosis, and reduced the expression of PI3K/AKT/mTOR proteins.50
In line with these observations, the current study investigated AKT and PI3K mRNA levels in AGS gastric cancer cells. The results showed that while the siRNA-mediated silencing of LINC00162 and cisplatin treatments separately decreased the expression of the AKT and PI3K genes, the expression of these genes was significantly lower in the combination treatment group compared to individual treatments and the control group. Furthermore, while LINC00162 silencing decreased the expression of these genes, the downregulation of PI3K was not significant.
Consistent with our findings, recent evidence further underscores the pivotal role of PI3K/Akt signaling in mediating drug resistance. Wang et al identified LINC00665 as a key oncogenic lncRNA contributing to trastuzumab resistance and tumorigenesis in gastric cancer via the PI3K/Akt pathway. LINC00665 was shown to sponge miR-199b-5p, upregulate SERPINE1 expression, and increase AKT phosphorylation, ultimately activating the PI3K/Akt pathway and promoting drug resistance.51 Similarly, PICSAR has been reported to function as an oncogene in hepatocellular carcinoma (HCC) by acting as a molecular sponge for miR-588. This interaction leads to upregulation of EIF6 expression, which in turn activates the PI3K/AKT/mTOR signaling pathway.37 Collectively, these findings support that silencing LINC00162 may enhance the efficacy of cisplatin through suppression of the PI3K/Akt signaling cascade.
Conclusion
For the first time, our pioneering research explored the impact of silencing LINC00162 via siRNA in conjunction with cisplatin on AGS gastric cancer cells. Collectively, these findings suggest that LINC00162 may function as an oncogene, and its inhibition significantly increased the susceptibility of AGS cells to cisplatin treatment. Furthermore, the combination treatment appeared to decrease cancer cell viability, induce apoptosis, inhibit progression, and cause cell cycle arrest. The evaluation of gene expression related to these biological processes also confirmed these findings. While LINC00162 siRNA alone had no significant effect on the expression of the BAX, CASP3, and TP53, the combination of the LINC00162 silencing and cisplatin treatment significantly altered the expression of these genes. AGS gastric cancer cells showed a reduction in migration and colony formation ability following LINC00162 silencing. This effect was also observed in the cells treated with cisplatin and those receiving the combination treatment. These results were in line with qRT-PCR assay findings, which showed reduced expression of the migration gene, MMP-9 and stemness gene, NANOG. Furthermore, our study revealed the role of LINC00162 in gastric cancer progression by decreasing AKT expression. In conclusion, these findings suggest that combining of LINC00162 silencing with cisplatin may represent a promising strategy for gastric cancer intervention and could improve patient survival.
However, this research has some limitations that need to be investigated in future studies. The experiments were performed in a single cell line under in vitro conditions, which may not fully reflect the tumor heterogeneity of the patients. Moreover, analyzing the expression of the proteins with a Western blot assay may help better understand the involvement of various signaling pathways. Therefore, future studies should validate these findings in multiple cell lines and in vivo models, and further investigate the upstream and downstream signaling pathways associated with LINC00162 to define its therapeutic potential in gastric cancer better.
Research Highlights
What is the current knowledge?
-
Cisplatin is first-line chemotherapy for gastric cancer, but resistance limits its effectiveness.
-
Long non-coding RNAs (lncRNAs) regulate apoptosis, cell cycle, migration, and contribute to chemoresistance.
-
lncRNAs like HOTAIR and ROR are linked to Cisplatin resistance; LINC00162’s role in the sensitivity of gastric cancer cells to cisplatin was unknown.
-
The PI3K/AKT pathway and stemness factors like NANOG drive gastric cancer progression, metastasis, and drug resistance.
-
Novel therapeutic strategies are needed to overcome Cisplatin resistance and improve patient survival.
What is new here?
-
This is the first study showing silencing LINC00162 (PICSAR) enhances Cisplatin sensitivity in AGS gastric cancer cells.
-
LINC00162 silencing reduced Cisplatin IC50, promoted apoptosis, induced sub-G1 cell cycle arrest, and decreased viability.
-
Combination treatment markedly altered the expression of apoptosis (BAX, BCL2, CASP3, CASP9), stemness (NANOG), and survival/metastasis genes (MMP-9, AKT, PI3K).
-
Dual treatment suppressed migration and colony formation, downregulated MMP-9 and NANOG, and inhibited PI3K/AKT signaling.
-
Findings suggest LINC00162 acts as an oncogene and propose its silencing as a potential therapeutic target to improve Cisplatin response.
Competing Interests
The authors declare that they have no conflicts of interest.
Data Availability Statement
All data generated or analyzed during this study are included in this published article
Ethical Approval
Not applicable.
Acknowledgements
The authors are thankful for the support of the Immunology Research Center, Tabriz University of Medical Science and Department of Biology, Ah.C., Islamic Azad University.
References
- Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin 2021; 71:209-49. doi: 10.3322/caac.21660 [Crossref] [ Google Scholar]
- Li GZ, Doherty GM, Wang J. Surgical Management of Gastric Cancer: A Review. JAMA Surg 2022; 157:446-54. doi: 10.1001/jamasurg.2022.0182 [Crossref] [ Google Scholar]
- Liu-Burdowski J, Park J. Treatment of Early Gastric Cancer. Surg Clin North Am 2025; 105:55-63. doi: 10.1016/j.suc.2024.06.006 [Crossref] [ Google Scholar]
- Zeng Y, Lockhart AC, Jin RU. The preclinical discovery and development of zolbetuximab for the treatment of gastric cancer. Expert Opin Drug Discov 2024; 19:873-86. doi: 10.1080/17460441.2024.2370332 [Crossref] [ Google Scholar]
- Wang G, Huang Y, Zhou L, Yang H, Lin H, Zhou S. Immunotherapy and targeted therapy as first-line treatment for advanced gastric cancer. Crit Rev Oncol Hematol 2024; 198:104197. doi: 10.1016/j.critrevonc.2023.104197 [Crossref] [ Google Scholar]
- Kang YK, Kang WK, Shin DB, Chen J, Xiong J, Wang J. Capecitabine/cisplatin versus 5-fluorouracil/cisplatin as first-line therapy in patients with advanced gastric cancer: a randomised phase III noninferiority trial. Ann Oncol 2009; 20:666-73. doi: 10.1093/annonc/mdn717 [Crossref] [ Google Scholar]
- Yan H, Cai X, Fu S, Zhang X, Zhang J. PRDX3 promotes resistance to cisplatin in gastric cancer cells. J Cancer Res Ther 2022; 18:1994-2000. doi: 10.4103/jcrt.jcrt_970_22 [Crossref] [ Google Scholar]
- Huang W, Wen F, Yang P, Li Y, Li Q, Shu P. Yi-qi-hua-yu-jie-du decoction induces ferroptosis in cisplatin-resistant gastric cancer via the AKT/GSK3β/NRF2/GPX4 axis. Phytomedicine 2024; 123:155220. doi: 10.1016/j.phymed.2023.155220 [Crossref] [ Google Scholar]
- Shang Z, Luo Z, Wang Y, Liu Q, Xin Y, Zhang M. CircHIPK3 contributes to cisplatin resistance in gastric cancer by blocking autophagy-dependent ferroptosis. J Cell Physiol 2023; 238:2407-24. doi: 10.1002/jcp.31093 [Crossref] [ Google Scholar]
- Esteller M, Pandolfi PP. The Epitranscriptome of Noncoding RNAs in Cancer. Cancer Discov 2017; 7:359-68. doi: 10.1158/2159-8290.Cd-16-1292 [Crossref] [ Google Scholar]
- Kapranov P, Cheng J, Dike S, Nix DA, Duttagupta R, Willingham AT. RNA maps reveal new RNA classes and a possible function for pervasive transcription. Science 2007; 316:1484-8. doi: 10.1126/science.1138341 [Crossref] [ Google Scholar]
- Gao N, Li Y, Li J, Gao Z, Yang Z, Li Y. Long Non-Coding RNAs: The Regulatory Mechanisms, Research Strategies, and Future Directions in Cancers. Front Oncol 2020; 10:598817. doi: 10.3389/fonc.2020.598817 [Crossref] [ Google Scholar]
- Li S, Lu C, Li X, Li F, Zhao Y, Xu M. LncRNA HOXA10-AS functions as an oncogene by binding miR-6509-5p to upregulate Y-box binding protein 1 in gastric cancer. Bioengineered 2022; 13:11373-87. doi: 10.1080/21655979.2022.2059615 [Crossref] [ Google Scholar]
- Li S, Sun J, Ma J, Zhou C, Yang X, Zhang S. LncRNA LENGA acts as a tumor suppressor in gastric cancer through BRD7/TP53 signaling. Cell Mol Life Sci 2022; 80:5. doi: 10.1007/s00018-022-04642-2 [Crossref] [ Google Scholar]
- Alamdari SG, Mohammadzadeh R, Amini M, Najafi S, Baradaran B, Bahojb Mahdavi SZ. Improvement of carboplatin chemosensitivity in lung cancer cells by siRNA-mediated downregulation of DLGAP1-AS2 expression. Sci Rep 2025; 15:7971. doi: 10.1038/s41598-025-87649-6 [Crossref] [ Google Scholar]
- Luo Y, Zheng S, Wu Q, Wu J, Zhou R, Wang C. Long noncoding RNA (lncRNA) EIF3J-DT induces chemoresistance of gastric cancer via autophagy activation. Autophagy 2021; 17:4083-101. doi: 10.1080/15548627.2021.1901204 [Crossref] [ Google Scholar]
- Akhgari H, Shokri N, Dehghanzadeh P, Tayefeh-Gholami S, Rajabi A, Safaralizadeh R. Expression pattern of PCAT1, PCAT2, and PCAT5 lncRNAs and their value as diagnostic biomarkers in patients with gastric cancer. Pathol Res Pract 2023; 248:154654. doi: 10.1016/j.prp.2023.154654 [Crossref] [ Google Scholar]
- Lu X, Gan Q, Gan C, Zheng Y, Cai B, Li X. Long non-coding RNA PICSAR knockdown inhibits the progression of cutaneous squamous cell carcinoma by regulating miR-125b/YAP1 axis. Life Sci 2021; 274:118303. doi: 10.1016/j.lfs.2020.118303 [Crossref] [ Google Scholar]
- Wang X, Zhang R, Wu S, Shen L, Ke M, Ouyang Y. Super-Enhancer LncRNA LINC00162 Promotes Progression of Bladder Cancer. iScience 2020; 23:101857. doi: 10.1016/j.isci.2020.101857 [Crossref] [ Google Scholar]
- Zong L, Hattori N, Yasukawa Y, Kimura K, Mori A, Seto Y. LINC00162 confers sensitivity to 5-Aza-2'-deoxycytidine via modulation of an RNA splicing protein, HNRNPH1. Oncogene 2019; 38:5281-93. doi: 10.1038/s41388-019-0792-8 [Crossref] [ Google Scholar]
- Hejazi M, Jafari T, Yari A, Heshmat R, Larijani B, Ahvaz S. A novel long non-coding RNA, PICSAR, promotes thyroid cancer progression through the hsa-miR-320A/hsa-miR-485/RAPGEFL1 axis. Med Oncol 2025; 42:448. doi: 10.1007/s12032-025-02987-9 [Crossref] [ Google Scholar]
- Wang D, Zhou X, Yin J, Zhou Y. Lnc-PICSAR contributes to cisplatin resistance by miR-485-5p/REV3L axis in cutaneous squamous cell carcinoma. Open Life Sci 2020; 15:488-500. doi: 10.1515/biol-2020-0049 [Crossref] [ Google Scholar]
- Xia JY, Aadam AA. Advances in screening and detection of gastric cancer. J Surg Oncol 2022; 125:1104-9. doi: 10.1002/jso.26844 [Crossref] [ Google Scholar]
- Patel TH, Cecchini M. Targeted Therapies in Advanced Gastric Cancer. Curr Treat Options Oncol 2020; 21:70. doi: 10.1007/s11864-020-00774-4 [Crossref] [ Google Scholar]
- Song Z, Wu Y, Yang J, Yang D, Fang X. Progress in the treatment of advanced gastric cancer. Tumour Biol 2017; 39:1010428317714626. doi: 10.1177/1010428317714626 [Crossref] [ Google Scholar]
- Wu C, Wang S, Huang T, Xi X, Xu L, Wang J. NPR1 promotes cisplatin resistance by inhibiting PARL-mediated mitophagy-dependent ferroptosis in gastric cancer. Cell Biol Toxicol 2024; 40:93. doi: 10.1007/s10565-024-09931-z [Crossref] [ Google Scholar]
-
Basu A, Krishnamurthy S. Cellular responses to Cisplatin-induced DNA damage. J Nucleic Acids 2010; 2010. doi: 10.4061/2010/201367.
- Marin JJ, Al-Abdulla R, Lozano E, Briz O, Bujanda L, Banales JM. Mechanisms of Resistance to Chemotherapy in Gastric Cancer. Anticancer Agents Med Chem 2016; 16:318-34. doi: 10.2174/1871520615666150803125121 [Crossref] [ Google Scholar]
- Azadi SS, Safaralizadeh R, Amini M, Hosseinpour Feizi MA, Abdolzadeh S, Najafi S. Investigating the effect of LncRNA DLGAP1-AS2 suppression on chemosensitivity of gastric cancer to chemotherapy. NaunynSchmiedebergs Arch Pharmacol 2024; 397:7891-903. doi: 10.1007/s00210-024-03130-7 [Crossref] [ Google Scholar]
- Chen Z, Zhang X, Li Z, Zhang H, Wang Z. lncRNA LINC02323 predicts adverse neoadjuvant chemotherapy outcomes of gastric cancer patients and regulates cell sensitivity to 5-fluorouracil by negatively modulating miR-139-3p. Ann Med 2024; 56:2424513. doi: 10.1080/07853890.2024.2424513 [Crossref] [ Google Scholar]
- Xiang W, Zhang B, Li H. LncRNA DLEU2 contributes to Taxol resistance of gastric cancer cells through regulating the miR-30c-5p-LDHA axis. J Chemother 2024; 36:49-60. doi: 10.1080/1120009x.2023.2203606 [Crossref] [ Google Scholar]
- Wang S, Chen W, Yu H, Song Z, Li Q, Shen X. lncRNA ROR Promotes Gastric Cancer Drug Resistance. Cancer Control 2020; 27:1073274820904694. doi: 10.1177/1073274820904694 [Crossref] [ Google Scholar]
- Yan J, Dang Y, Liu S, Zhang Y, Zhang G. LncRNA HOTAIR promotes cisplatin resistance in gastric cancer by targeting miR-126 to activate the PI3K/AKT/MRP1 genes. Tumour Biol 2016; 37:16345-55. doi: 10.1007/s13277-016-5448-5 [Crossref] [ Google Scholar]
- Hejazi M, Bahojb Mahdavi SZ, Abedimanesh S, Heshmat R, Larijani B, Mokhtarzadeh AA. LINC00162 silencing enhances sorafenib sensitivity and inhibits thyroid cancer cells progression through modulation of MAPK signaling and apoptosis. Sci Rep 2025; 15:29726. doi: 10.1038/s41598-025-12805-x [Crossref] [ Google Scholar]
- Qian S, Wei Z, Yang W, Huang J, Yang Y, Wang J. The role of BCL-2 family proteins in regulating apoptosis and cancer therapy. Front Oncol 2022; 12:985363. doi: 10.3389/fonc.2022.985363 [Crossref] [ Google Scholar]
- Huang KH, Fang WL, Li AF, Liang PH, Wu CW, Shyr YM. Caspase-3, a key apoptotic protein, as a prognostic marker in gastric cancer after curative surgery. Int J Surg 2018; 52:258-63. doi: 10.1016/j.ijsu.2018.02.055 [Crossref] [ Google Scholar]
- Liu Z, Mo H, Sun L, Wang L, Chen T, Yao B. Long noncoding RNA PICSAR/miR-588/EIF6 axis regulates tumorigenesis of hepatocellular carcinoma by activating PI3K/AKT/mTOR signaling pathway. Cancer Sci 2020; 111:4118-28. doi: 10.1111/cas.14631 [Crossref] [ Google Scholar]
- Lee WJ, Ji H, Jeong SD, Pandey PR, Gorospe M, Kim HH. LINC00162 regulates cell proliferation and apoptosis by sponging PAQR4-targeting miR-485-5p. J Cell Physiol 2022; 237:2943-60. doi: 10.1002/jcp.30758 [Crossref] [ Google Scholar]
- Iv Santaliz-Ruiz LE, Xie X, Old M, Teknos TN, Pan Q. Emerging role of nanog in tumorigenesis and cancer stem cells. Int J Cancer 2014; 135:2741-8. doi: 10.1002/ijc.28690 [Crossref] [ Google Scholar]
- Zhang J, Wang X, Chen B, Xiao Z, Li W, Lu Y. The human pluripotency gene NANOG/NANOGP8 is expressed in gastric cancer and associated with tumor development. Oncol Lett 2010; 1:457-63. doi: 10.3892/ol_00000081 [Crossref] [ Google Scholar]
- Basati G, Mohammadpour H, Emami Razavi A. Association of High Expression Levels of SOX2, NANOG, and OCT4 in Gastric Cancer Tumor Tissues with Progression and Poor Prognosis. J Gastrointest Cancer 2020; 51:41-7. doi: 10.1007/s12029-018-00200-x [Crossref] [ Google Scholar]
- Vasefifar P, Najafi S, Motafakkerazad R, Amini M, Safaei S, Najafzadeh B. Targeting Nanog expression increased Cisplatin chemosensitivity and inhibited cell migration in Gastric cancer cells. Exp Cell Res 2023; 429:113681. doi: 10.1016/j.yexcr.2023.113681 [Crossref] [ Google Scholar]
- Ohtsuka J, Oshima H, Ezawa I, Abe R, Oshima M, Ohki R. Functional loss of p53 cooperates with the in vivo microenvironment to promote malignant progression of gastric cancers. Sci Rep 2018; 8:2291. doi: 10.1038/s41598-018-20572-1 [Crossref] [ Google Scholar]
- Scheau C, Badarau IA, Costache R, Caruntu C, Mihai GL, Didilescu AC. The Role of Matrix Metalloproteinases in the Epithelial-Mesenchymal Transition of Hepatocellular Carcinoma. Anal Cell Pathol (Amst) 2019; 2019:9423907. doi: 10.1155/2019/9423907 [Crossref] [ Google Scholar]
- Liu L, Ye Y, Zhu X. MMP-9 secreted by tumor associated macrophages promoted gastric cancer metastasis through a PI3K/AKT/Snail pathway. Biomed Pharmacother 2019; 117:109096. doi: 10.1016/j.biopha.2019.109096 [Crossref] [ Google Scholar]
- Piipponen M, Nissinen L, Farshchian M, Riihilä P, Kivisaari A, Kallajoki M. Long Noncoding RNA PICSAR Promotes Growth of Cutaneous Squamous Cell Carcinoma by Regulating ERK1/2 Activity. J Invest Dermatol 2016; 136:1701-10. doi: 10.1016/j.jid.2016.03.028 [Crossref] [ Google Scholar]
- Luo M, Liang C. LncRNA LINC00483promotes gastric cancer development through regulating MAPK1 expression by sponging miR-490-3p. Biol Res 2020; 53:14. doi: 10.1186/s40659-020-00283-6 [Crossref] [ Google Scholar]
- Matsuoka T, Yashiro M. The Role of PI3K/Akt/mTOR Signaling in Gastric Carcinoma. Cancers (Basel) 2014; 6:1441-63. doi: 10.3390/cancers6031441 [Crossref] [ Google Scholar]
- Tao K, Yin Y, Shen Q, Chen Y, Li R, Chang W. Akt inhibitor MK-2206 enhances the effect of cisplatin in gastric cancer cells. Biomed Rep 2016; 4:365-8. doi: 10.3892/br.2016.594 [Crossref] [ Google Scholar]
- Wu Q, Zhu C, Zhao T, Liu T, Da M. Downregulation of LncRNA CCAT1 Enhances Chemosensitivity in Cisplatin-Resistant Gastric Cancer Cells. Drug Dev Res 2025; 86:e70048. doi: 10.1002/ddr.70048 [Crossref] [ Google Scholar]
- Wang B, Liu W, Song B, Li Y, Wang Y, Tan B. Targeting LINC00665/miR-199b-5p/SERPINE1 axis to inhibit trastuzumab resistance and tumorigenesis of gastric cancer via PI3K/AKt pathway. Noncoding RNA Res 2025; 10:153-62. doi: 10.1016/j.ncrna.2024.07.004 [Crossref] [ Google Scholar]