Bioimpacts. 16:32903.
doi: 10.34172/bi.32903
Review
The involvement of circular RNAs in breast cancer metastasis: Insight into EMT-related signaling pathways
Mehran Molavand Investigation, Writing – original draft, 1, 2, * 
Maryam Majidinia Investigation, Writing – review & editing, 3
Bahman Yousefi Conceptualization, Supervision, Writing – review & editing, 1, *
Author information:
1Department of Clinical Biochemistry and Laboratory Medicine, Faculty of Medicine, Tabriz University of Medical Sciences, Tabriz, Iran
2Student Research Committee, Tabriz University of Medical Sciences, Tabriz, Iran
3Solid Tumor Research Center, Cellular and Molecular Medicine Institute, Urmia University of Medical Sciences, Urmia, Iran
Abstract
Breast cancer, a complex and heterogeneous disease marked by uncontrolled cell growth in breast tissue, presents a significant global health challenge due to its high mortality rate. Metastasis, a major driver of cancer progression and mortality, poses a significant challenge in clinical oncology. This complex process involves profound changes in cytoskeletal dynamics and interactions with the extracellular matrix, including integrin-mediated adhesion and the activity of matrix metalloproteinases (MMPs). Circular RNAs (circRNAs) are a type of endogenous non-coding RNA with a covalently closed-loop structure. They serve as critical regulators of gene expression and modulate essential cellular processes, including proliferation, cell cycle control, and invasion. Through intricate interactions with key signaling pathways, including Wnt/β-catenin, PI3K/AKT, TGF-β/SMAD, and the MEK-MAPK pathway, circRNAs modulate these processes, particularly epithelial-to-mesenchymal transition (EMT), which is crucial in breast cancer metastasis. Mechanistically, circRNAs activate transcription factors associated with EMT, such as Slug, Snail, Twist, and Zeb. Moreover, circRNAs have emerged as promising clinical biomarkers in breast cancer, offering new opportunities for early diagnosis, prognostic assessment, and the development of targeted therapeutic strategies. In this article, the role of circRNAs in modulating EMT-regulating signaling pathways in the context of breast cancer metastasis will be reviewed.
Graphical Abstract
Keywords: CircRNAs, Breast cancer, EMT, Signaling pathways, Metastasis
Copyright and License Information
© 2026 The Author(s).
This work is published by BioImpacts as an open access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (
http://creativecommons.org/licenses/by-nc/4.0/). Non-commercial uses of the work are permitted, provided the original work is properly cited.
Funding Statement
Not applicable.
Introduction
Significance
The importance of studying circRNAs lies in the fact that they are in close contact with the regulation of various biological processes such as proliferation, gene expression, metastasis, and can also be indicators of pathological conditions. Investigating the circRNA profile in patients with metastatic breast cancer is seen as the initial crucial step towards leveraging circRNAs to combat breast cancer and its metastatic progression, ultimately enhancing prognosis through targeted therapeutic interventions.
Worldwide, breast cancer is the 5th leading cause of cancer-related death and the most prominent cause of cancer-related death in women. As a highly heterogeneous disease, understanding its tissue origin and classification is critical for effective diagnosis and treatment. The breast is anatomically composed of glandular, stromal, and lymphatic tissues.1,2 The glandular tissue is primarily responsible for the synthesis and secretion of milk. The cells most commonly involved in breast cancer originate from the glandular tissue, which consists of ducts (facilitating milk transport) and lobules (functioning in milk production).2 To guide clinical management and research, breast cancer is classified based on both histopathological and molecular features. Histologically, lobular carcinoma in situ (LCIS) and ductal carcinoma in situ (DCIS) are the two main types of non-invasive breast cancer, and infiltrating (or invasive) ductal carcinoma (IDC) and infiltrative lobular carcinoma (ILC) constitute the two main invasive types of breast cancer. It should be noted that IDC is responsible for about 80% of breast cancer's total incidence, while ILC is responsible for 10-15% of it. Molecular subtyping, which is determined by the expression of estrogen receptor (ER), progesterone receptor (PR), and HER2, further stratifies tumors into luminal A, luminal B, HER2-positive, and triple-negative subtypes (Table 1).3-5
Table 1.
Molecular subtypes of breast cancer
|
Molecular Subtype
|
Receptor Status
|
Prognosis
|
Incidence (%)
|
Treatment
|
Common Metastatic Site
|
Ref.
|
| Triple-negative (basal-like) |
ER-, PR-, HER2- |
Poor |
15-20 |
Targeted therapy |
Lung, Brain, Bone |
6,7
|
| HER2 + |
ER-, PR-, HER2 + |
Variable |
10-15 |
Targeted therapy |
Liver, Brain |
8,9
|
| Luminal A |
ER + , PR + , HER2- |
Good |
40 |
Endocrine and Targeted therapy |
Bone, Liver |
10,11
|
| Luminal B |
ER + , PR + , HER2 + /- |
Variable |
20 |
Endocrine and Targeted therapy |
Bone, Lung, Liver |
12,13
|
Studies have reported the risk factors involved in breast cancer in the form of two categories. The first category is assigned to non-manipulable conditions such as genetic background, mutation, aging, and race. On the contrary, the second category refers to conditions that a person has the ability to improve or eliminate, such as alcohol consumption, smoking, obesity, drugs, and chemicals.14,15 By integrating anatomical, histological, molecular, and epidemiological perspectives, we set the stage to explore how dysregulated regulatory networks drive breast cancer metastasis.
The metastatic cascade in cancer involves multiple sequential stages, beginning with cellular changes that promote increased motility and detachment from the primary tumor. Subsequent phases include molecular adaptations that enable cancer cells to intravasate into, survive within, and extravasate from the circulatory system, thereby facilitating colonization of distant organs.16,17 In breast cancer, epithelial-mesenchymal transition (EMT) is recognized as a critical early event in metastasis, characterized by the loss of epithelial polarity and intercellular adhesion, coupled with the acquisition of migratory and invasive mesenchymal traits.16 Multiple signaling axes, including TGF-β/SMAD, Wnt/β-catenin, PI3K/AKT, and MEK/MAPK, converge on EMT-associated transcription factors such as Snail, Slug, and Twist.18 Elucidating the signaling-based molecular regulators that orchestrate these critical metastatic processes is essential for the development of novel and effective therapeutic strategies.
Circular RNAs (circRNAs) are a large class of long non-coding RNAs characterized by a covalently closed loop structure, lacking the typical 5' cap and 3' poly(A) tail found in linear RNAs. The covalently closed structure resulting from back-splicing endows these molecules with remarkable stability.19 Functionally, many circRNAs act as molecular sponges for microRNAs (miRNAs), thereby modulating gene expression post-transcriptionally. In addition, they can interact with RNA-binding proteins, serve as scaffolds for protein complexes, or, in some cases, be translated into peptides, expanding their functional repertoire. Accordingly, their involvement in key processes like proliferation, gene expression, and metastasis makes circRNAs potential indicators of pathological conditions.20 Among these pathological processes, metastasis in breast cancer remains the leading cause of mortality, making it a critical focus for exploring the regulatory roles of circRNAs. Given EMT’s pivotal role in cancer cell dissemination, understanding its circRNA-mediated control offers valuable insight into breast cancer metastatic progression. In this review, relevant studies were gathered from major databases without language restrictions to clarify how circRNAs contribute to breast cancer metastasis, particularly through their involvement in EMT and associated signaling pathways.
Breast cancer pathogenesis
The development and progression of breast cancer are influenced by three major factors: hormonal, genetic, and environmental. These factors collectively contribute to breast tumorigenesis by promoting cellular proliferation and suppressing apoptosis.
(i) Hormonal factors: Estrogen and progesterone are the physiological triggers of the development of the female reproductive system through their receptors, ER and PR, respectively.21 Interestingly, ER and PR expression is restricted to a subset of terminally differentiated breast epithelial cells, which are non-proliferative. Therefore, most of the breast cells are not sensitive to these hormones and divide independently of ER and PR. This small population, in response to estrogen and progesterone, by secreting mediators such as Wnt and insulin-like growth factor 2, triggers the development and proliferation of adjacent cells. Unlike normal breast cells, the majority of breast cancer cells are ER and PR positive. Consequently, estrogen and progesterone play a pivotal role in promoting breast cancer cell growth.22,23 In addition to the oncogenic role, some studies have partially revealed a tumor-suppressive role for certain estrogen receptor isoforms, which can be valuable in the handling of breast cancer.24
(ii) Environmental factors: Metalloestrogens are divalent metals, such as cadmium, copper, zinc, iron, nickel, aluminum, arsenic, cobalt, and tin, that can mimic estrogenic activity by binding to the hormone-binding domain of estrogen receptor alpha (ERα). Therefore, by stimulating the expression of the target genes of estrogen, they can stimulate the proliferation and development of breast cancer.25,26 A large body of studies has acknowledged organochlorine pesticides, solar ultraviolet radiation, and ionizing radiation as environmental factors associated with breast cancer. Even though the escalation in the potential of DNA damage and the molecular level of reactive oxygen species (ROS), because of the mentioned factors, are the first-hand candidates for the connection with breast cancer, in cases such as organochlorine pesticides, the mechanism that increases the risk of breast cancer is still unknown.27-29
(iii) Genetic factors: Permanent genetic alterations, including deletions and mutations, increase the risk of breast cancer. The studies conducted to determine the involvement of genetic changes in the overall incidence of breast cancer show that the cause of 5-10% of breast cancer is due to hereditary abnormalities, and 90% is because of acquired changes in genetics.30 BRCA1 and BRCA2 genes, which are implicated in various processes such as DNA repair, are known as the most likely breast cancer predisposing genes that increase the risk of breast cancer by 11.4-fold and 11.7-fold, respectively.31 Studies have shown that mutations in BRCA1 and BRCA2 account for nearly 50% of hereditary breast cancer cases.32 In addition, TP53, PTEN, STK11, CDH1, PALB2, CHEK2, and CHEK2 are also among the most known predisposing genes.33
Molecular mechanisms of breast cancer metastasis; from EMT to colonization
The final stage in the evolutionary process of many malignancies is the metastasis-inducing phase, which is directly associated with poor prognosis. In breast cancer, this phase involves the formation of micrometastases at secondary sites, primarily including bone (up to 60%), lung (up to 34%), liver (up to 20%), and brain (up to 16%). However, with advances in early detection, effective screening methods such as mammography have gained clinical significance, leading to a significant reduction in breast cancer–related mortality in recent years.34-37
From a molecular perspective, the metastatic process can be categorized into three phases. The first phase involves the triggering of molecular alterations and the enhancement of dissemination capacity into the circulatory system, primarily characterized by EMT.16
EMT is a multistep dedifferentiation process, during which cells acquire a hybrid state expressing both epithelial and mesenchymal markers. This process can be divided into three types: the first primarily functions in embryonic development, the second contributes to wound healing and tissue regeneration, and the third is associated with cancer progression by enhancing metastatic potential.38
The activation of EMT-associated transcription factors (e.g., Slug, Snail, ZEB, and Twist) drives a cascade of cellular alterations, the cumulative effect of which culminates in enhanced motility and invasiveness. In more detail, the downregulation of adhesion molecules such as E-cadherin, occludin, claudins, desmoplakin, EpCAM, and plakophilin, alongside the upregulation of mesenchymal markers associated with cellular motility, including N-cadherin, vimentin, fibronectin, integrin β6, and α-SMA, represents a prominent cellular strategy for detachment from the surrounding matrix.39,40 In parallel, the induction of cytoskeletal remodeling and the activation of matrix metalloproteinases (MMPs) such as MMP15, MMP2, and MMP9 further potentiate this migratory and invasive phenotype.39,41 It is noteworthy that integrin, as an adhesion receptor implicated in tissue integrity, activates MMP.42
However, the primary challenge associated with cellular detachment during EMT is the induction of programmed cell death triggered by loss of contact with the extracellular matrix, known as anoikis.43 Circumventing anoikis involves multiple mechanisms, among which the upregulation of anti-apoptotic components such as survivin, Bcl-2, and Bcl-xL, along with the activation of Focal Adhesion Kinase (FAK)-dependent survival pathways, are the most well-established.16 Nevertheless, studies have also documented the roles of Twist and ZEB1 in enhancing cell survival potential during EMT through mechanisms involving the suppression of p14ARF and p73, respectively.44
The second phase pertains to the plasticity of circulatory tumor cells (CTCs). Phenotypic plasticity in CTCs allows them to dynamically oscillate between epithelial and mesenchymal states. This plasticity is controlled by signaling pathways such as TGF-β, hypoxia-induced HIF-1α, Wnt/β-catenin, Notch, and Hedgehog, which in turn activate EMT transcription factors including Snail, Slug, Twist, and ZEB1/2.16 As a result, CTCs often acquire a hybrid epithelial/mesenchymal phenotype that confers enhanced survival and motility, as well as stem-like traits such as self-renewal and unlimited proliferative capacity.45
A cancer cell that enters the bloodstream implements tactics to escape from life-threatening factors, such as the immune system and apoptosis, induced by the lack of attachment to the matrix. One of them is activation and attachment to platelets, respectively, through the augmented expression of tissue factor and P-selectin ligands.46 Furthermore, platelets secrete TGF-β, which promotes the epithelial-mesenchymal transition (EMT). They also physically shield tumor cells from NK cell-mediated lysis. In this regard, studies have shown that increased platelet count is associated with poor prognosis in malignancies.47
The third phase involves the extravasation of CTCs to colonize and establish microscopic tumors at secondary sites. This phase hinges on a finely tuned interplay among CTCs, endothelial cells, and extracellular matrix (ECM) components, in which adhesion molecules (selectins, ICAM, and VCAM) initiate vascular arrest and MMPs orchestrate basement membrane degradation and tissue invasion.48,49 Effective colonization then demands tumor cell adaptation to the new microenvironment, immune evasion, and neoangiogenesis. At this stage, phenotypic plasticity marked by a partial return to epithelial traits is important for successful engraftment.50 Notably, some disseminated cells may enter dormancy, retaining the capacity for later reactivation and secondary lesion formation.51
A concise overview of this cascade clearly highlights the critical importance of the EMT process; therefore, focusing on this aspect could represent a significant step forward in overcoming metastasis in breast cancer.
Circular RNAs: biogenesis and methodological landscape
Formation of circRNAs from pre-mRNAs predominately occurs by back splicing or exon skipping. The closed-loop form of this class of non-coding RNAs provides durability against exonucleases due to the absence of free ends. This means that the 3' and 5' ends are linked via covalent bonds.20
Currently, there is significant interest in three models that elucidate the formation process of circRNAs. Model 1: RNA binding protein (RBP)-mediated circularization, which is based on the regulatory effect of various RBPs, such as DHX9, QKI, and MBL, on adjoining splicing sites. Model2: Circularization based on intron pairing, which is based on the pairing of reverse complementary sequences, especially Alu elements, in adjacent introns. Model3: Exon skipping or lariat-based circularization, which is based on the formation of a lariat with RNA splicing. This process can be explained by joining the 3' end of one exon with the 5' end of another exon and separating the intron and exon between these two exons in a circular form (Fig. 1).52,53
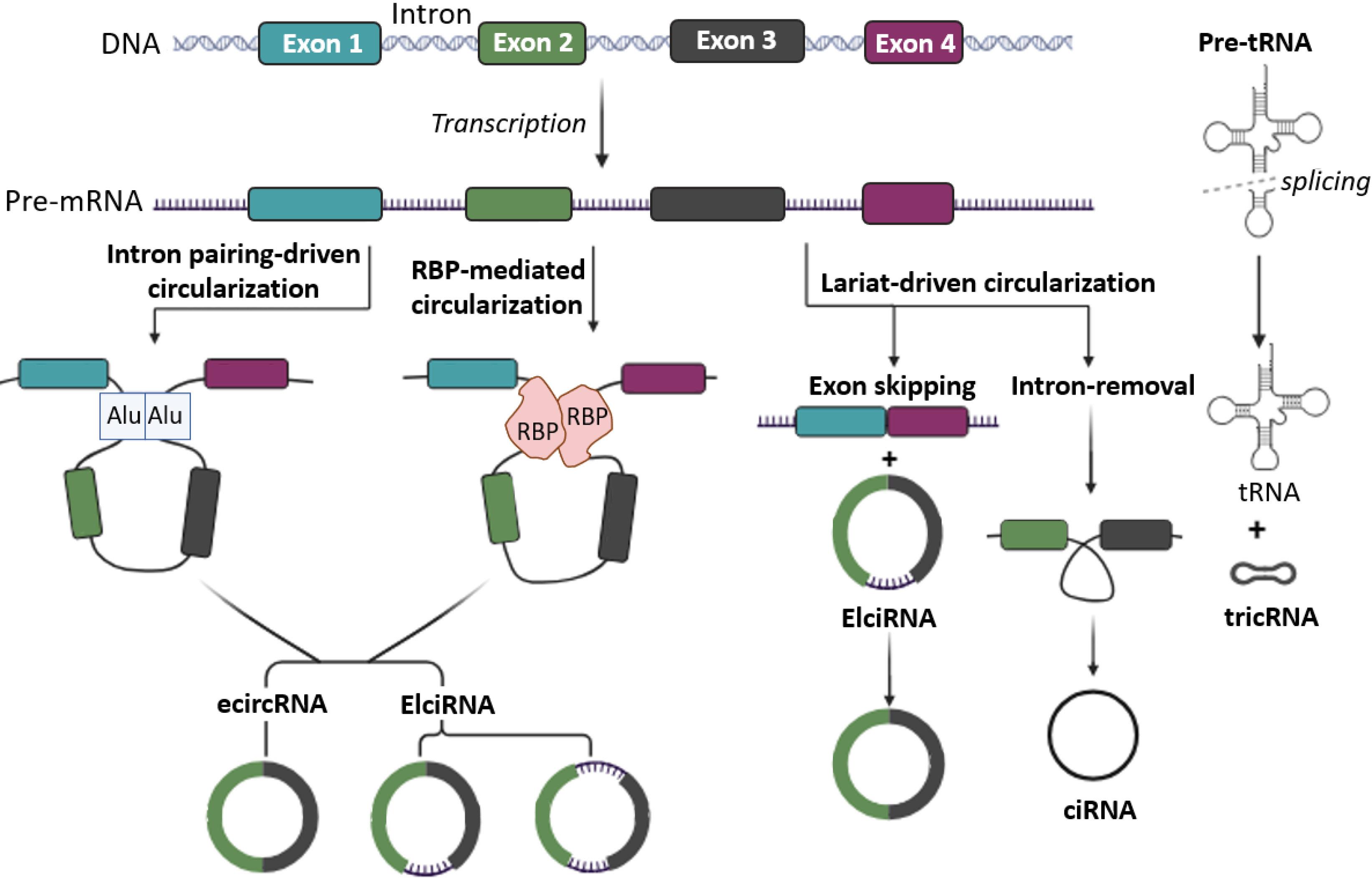
Fig. 1.
Schematic illustration of the core models presented in the construction of circRNA. Circularization based on intron pairing, RNA binding protein (RBP) mediated circularization and Exon skipping or lariat-based circularization. The circRNA family encompasses four distinct structural groups: exonic circular RNA (ecircRNA), circular intronic RNA (ciRNA), exon-intron circular RNA (EIciRNA), and tRNA-intronic circular RNA (tricRNA).
.
Schematic illustration of the core models presented in the construction of circRNA. Circularization based on intron pairing, RNA binding protein (RBP) mediated circularization and Exon skipping or lariat-based circularization. The circRNA family encompasses four distinct structural groups: exonic circular RNA (ecircRNA), circular intronic RNA (ciRNA), exon-intron circular RNA (EIciRNA), and tRNA-intronic circular RNA (tricRNA).
The categorization of circRNAs into four distinct groups based on their constituent parts has opened avenues for deeper investigation. In addition, the names of the four groups are clearly related to their components, including (i) exonic circular RNA or ecircRNA, (ii) circular intronic RNA or ciRNA, (iii) exon-intron circular RNA or EIciRNA, and (iv) tRNA-intronic circular RNA or tricRNA. For instance, ecircRNAs consist solely of exonic sequences, whereas ciRNAs are composed of intronic sequences. EIciRNAs contain both exonic and intronic regions, while tricRNAs contain tRNA sequences within their structure. This clear categorization facilitates targeted investigations into the roles and regulatory mechanisms of circRNAs in various biological processes and diseases (Fig. 1).54-56
Noteworthy, circRNAs contribute to cellular stress responses by modulating key signaling pathways such as MAPK, p53, and NF-κB.57,58 They are also detected in stress granules, membrane-less ribonucleoprotein structures that help cells adapt to conditions like nutrient deprivation, oxidative stress, heat shock, and viral infection.59 Stress granules temporarily halt translation and protect mRNAs. While the presence of circRNAs in stress granules suggests their involvement in stress responses, much about their specific role remains unclear.60
Studies in this area typically start with whole-transcriptome RNA sequencing (RNA-seq) paired with specialized bioinformatic pipelines to detect backsplice junctions characteristic of circRNAs.61 Because circRNAs constitute only a minor fraction of total RNA, samples are usually enriched by depleting ribosomal RNA and selecting against polyadenylated transcripts, often combining both approaches to maximize circRNA recovery.62,63 In some workflows, Rnase R treatment is also applied to selectively degrade linear RNAs, further enhancing the relative abundance of circular species.61 Definitive validation hinges on PCR amplification of the backsplice junction using divergent primers: each primer anneals just outside the junction and faces away from its partner, yielding a product only if the template is truly circular.64 Ultimately, northern blotting remains the gold standard for directly visualizing circRNA size and topology.65
Several methodological challenges complicate circRNA detection. For instance, backsplice junction reads from RNA-seq may result from repeated exonic regions or overlapping transcripts rather than true circular RNAs. Moreover, reverse transcriptase can sometimes switch templates during cDNA synthesis, creating chimeric artifacts that resemble circular junctions in PCR. It should be noted that circRNA prediction algorithms vary considerably in their stringency and detection criteria. Studies have shown that between 10% and 45% of predicted circRNAs may represent false positives, and approximately 40% of identified candidates are reported by only a single detection tool.62,66
For future circRNA research, it is essential to integrate multiple detection algorithms with biochemical verification methods and to employ unique molecular identifiers to minimize artefacts.67-69 Moreover, adopting three-dimensional culture systems and single-cell RNA sequencing, together with comprehensive integration of TCGA datasets and nanopore sequencing data, will facilitate the direct translation of experimental findings into clinical applications.70-72
Role of circRNAs in metastatic cascade
A large body of studies has demonstrated the multifaceted involvement of circRNAs in the metastatic cascade of breast cancer. In light of the critical role of EMT, we delineate these interactions into two main domains: circRNA-mediated regulation of EMT and circRNA-driven control of other metastatic phases.
Circular RNAs in the regulation of early-stage; EMT
As previously stated, EMT is a multistep dedifferentiation process that converts polarized epithelial cells into non-polarized mesenchymal cells. Four general steps make up the EMT process in cancer cell metastasis: 1) initiation, 2) loss of epithelial properties, 3) gain of mesenchymal characteristics, and 4) remodeling of cytoskeleton and cell morphology (Fig. 2).
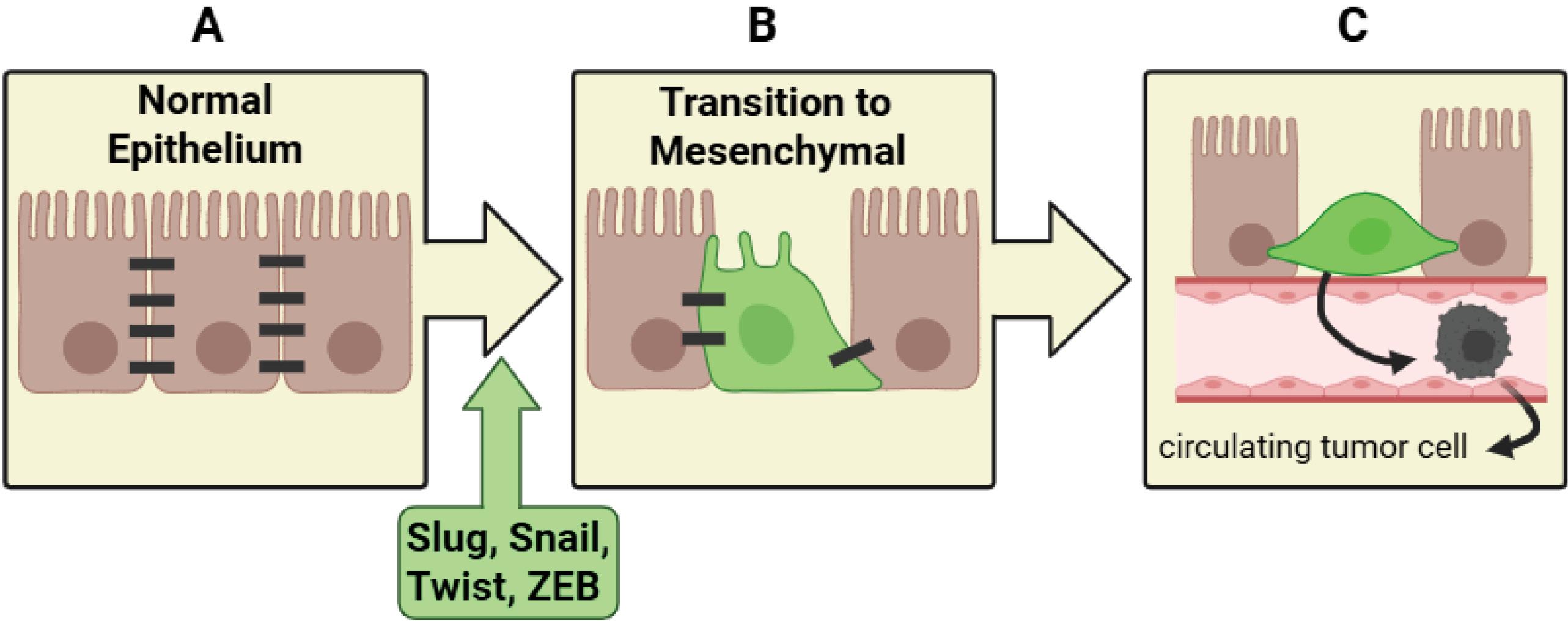
Fig. 2.
Schematic illustration of the EMT process. (A) Normal epithelium: Epithelial cells maintain robust intercellular adhesions through well-organized junctional complexes, preserving tissue architecture and polarity. (B) EMT induction: Upregulation of EMT-associated transcription factors, including Snail, Slug, Twist, and ZEB, leads to the repression of epithelial markers and activation of mesenchymal genes, resulting in morphological changes and enhanced cellular motility. (C) Invasion and intravasation: Mesenchymal-like tumor cells detach from the primary epithelium and enter the bloodstream, enabling them to infiltrate distant sites and establish metastatic colonies.
.
Schematic illustration of the EMT process. (A) Normal epithelium: Epithelial cells maintain robust intercellular adhesions through well-organized junctional complexes, preserving tissue architecture and polarity. (B) EMT induction: Upregulation of EMT-associated transcription factors, including Snail, Slug, Twist, and ZEB, leads to the repression of epithelial markers and activation of mesenchymal genes, resulting in morphological changes and enhanced cellular motility. (C) Invasion and intravasation: Mesenchymal-like tumor cells detach from the primary epithelium and enter the bloodstream, enabling them to infiltrate distant sites and establish metastatic colonies.
Transcription factors directly implicated in inhibiting CDH1 expression include the Snail family (Snail, Slug, Smuc), ZEB family (Zeb1/2), and b-HLH family (Twist1/2). Additionally, there are indirect repressors of CDH1, such as FoxC2, Goosecoid, TCF4, and PRRX1. These regulatory elements collectively drive the progression of EMT, facilitating the phenotype transformation.16
In the subsequent phase, crucial proteins responsible for maintaining cell-to-cell connections and apical-basal cell polarity, including those involved in tight junctions, adherens junctions, and gap junctions, are downregulated. In other words, proteins such as E-cadherin, cytokeratin, connexins, desmoplakin, zonula occludens, beta-catenin, and occludin experience systematic reduction during this phase of cellular transformation.16 The cell develops mesenchymal traits in the following stage. A closer look reveals that the cell exhibits a notable resistance to apoptosis and that its motility/invasive capacity is increased by the elevated expression of mesenchymal markers such as vimentin, fibronectin, N-cadherin, and MMPs.16,73
The three main components of the cytoskeleton are intermediate filaments, microtubules, and actin.74,75 Intermediate filaments, mainly composed of keratins and vimentin, provide structural support and flexibility to cells.76 Notably, EMT involves a switch from epithelial-specific keratins to mesenchymal-specific vimentin, a hallmark of this transition.77,78 Actin stress fibers, formed during EMT, offer contractile strength essential for cell movement and invasion.79,80 Meanwhile, the disorganized microtubule network in mesenchymal cells promotes the creation of membrane protrusions like filopodia and lamellipodia, aiding cell movement.81,82 These fundamental alterations in the cytoskeleton induce morphological changes, transitioning cells from their typical cuboidal epithelial shape to an elongated, spindle-like mesenchymal morphology.83 Overall, a cell gains the ability to separate from the primary site, enter and exit blood and lymphatic vessels, colonize the secondary site, and form microscopic metastatic tumors after passing through the transformational process of EMT.16 The intimate connection between signaling pathways and the molecular alterations driving EMT progression is readily apparent. Therefore, by focusing investigations on the circRNA-signaling effector axis, researchers can uncover novel regulatory mechanisms and identify potential therapeutic targets to modulate EMT.
TGFβ/SMAD mediated regulation of EMT
Both TGF-βRI and TGF-βRII possess serine/threonine kinase activity; however, TGF-βRII is constitutively active.84 Upon ligand (in dimer form) binding, TGF-βRI and TGF-βRII form a heteromeric receptor complex, leading to the activation of TGF-βRI through phosphorylation of its cytoplasmic domain by TGF-βRII. This phosphorylation event triggers the initiation of downstream signaling pathways. TGF-βRI phosphorylates the SSXS motif of receptor-regulated Smads (R-Smads), such as Smad2 and Smad3, in a process facilitated by SARA. This leads to the formation of R-Smad/Smad4 complexes. Subsequently, these complexes translocate into the nucleus to regulate gene expression. It's worth noting that Smads are mediators of the TGF-β family, and TGF-βRI phosphorylates both Smad2 and Smad3, members of the R-Smad family, in response to TGF-β signaling. After translocation into the nucleus, the R-Smad/Smad4 complex regulates gene expression. However, Smad7 (an inhibitory Smad) can suppress this pathway by blocking the activation of TGF-βRI.85-92 Finally, Smad can regulate the expression of EMT acceleration factors such as Slug, Zeb, Snail, and Twist (Fig. 3).93
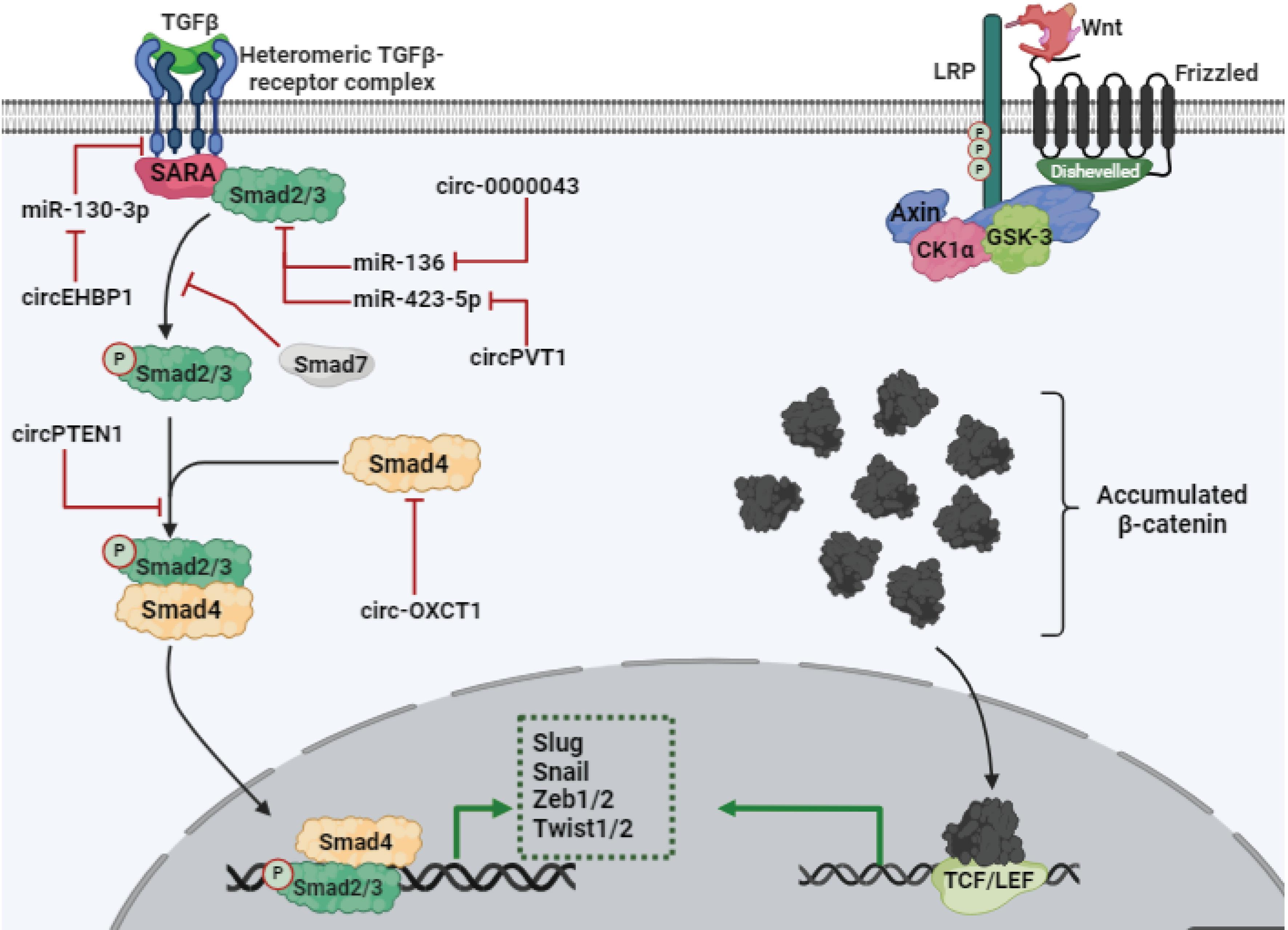
Fig. 3.
Schematic illustration of TGF-β/Smad and Wnt/β-catenin signaling pathways. Both pathways are related to EMT, mainly by the expression of transcription factors related to EMT (Slug, Zeb1/2, Snail, and Twist1/2).
.
Schematic illustration of TGF-β/Smad and Wnt/β-catenin signaling pathways. Both pathways are related to EMT, mainly by the expression of transcription factors related to EMT (Slug, Zeb1/2, Snail, and Twist1/2).
The research findings demonstrate a correlation between EMT and the TGF-β signaling pathway. Elevated levels of TGF-β, induced by the activation of the upstream stimulatory family 1 (USF1), have been associated with increased EMT. Notably, studies suggest that miR-148a-3p and miR-152-3p act as inhibitors of USF1, thereby regulating EMT. CircANKS1B, identified for its role in promoting malignant characteristics in breast cancer cells and exacerbating prognosis, indirectly boosts TGF-β-induced EMT by suppressing these two miRNAs.94,95 Consistent with these observations, circ-0000043, recognized for its oncogenic properties in breast cancer, exacerbates EMT by suppressing miR-136, through a Smad3-dependent mechanism.96
Although studies in this area remain limited in breast cancer, the circRNA/miRNA/TGFβ/Smad/EMT axis has been well characterized in other malignancies. Targeted investigations based on these findings may accelerate progress and enhance our understanding of its role in breast cancer metastasis. Several examples stand out clearly in this regard. In gastric cancer, the circRNAs circPVT1 and circOXCT1 exhibit opposing roles in EMT regulation through their interaction with the TGFβ signaling pathway. CircPVT1 acts as a sponge for miR-423-5p, which leads to the upregulation of Smad3, promoting EMT. Conversely, circ-OXCT1 inhibits EMT by attenuating the TGF-β pathway through suppressing Smad4.97,98 Similarly, research has highlighted the roles of circPTK2 (hsa_circ_0008305) and circEHBP1 in modulating EMT. CircPTK2 inhibits EMT in lung cancer by up-regulating TIF1γ through the inhibition of miR-429/miR-200b-3p, thus attenuating the TGF-β-induced EMT process. On the other hand, circEHBP1 enhances EMT in bladder cancer by up-regulating TGF-βR1 via the inhibition of miR-130-3p.99,100 Furthermore, in colorectal cancer, circPTEN1 inhibits TGF-β-induced EMT by preventing the binding of Smad4 to Smad2/3.101 Overall, despite the noticeable scarcity of related data, the critical role of circRNAs in the fine-tuned regulation of TGFβ/SMAD-driven EMT remains a reliable and compelling aspect of current evidence.
Wnt/β-catenin mediated regulation of EMT
In the Wnt/β-catenin signaling pathway, the control center revolves around the buildup of a crucial cytoplasmic co-activator called β-catenin. When Wnt signaling is inactive, a specialized degradation complex remains highly active, tasked with preventing the accumulation of β-catenin.102 This complex comprises key components: Axin, adenomatous polyposis coli (APC), glycogen synthase kinase 3 (GSK-3, a Ser/Thr kinase), and casein kinase 1α (CK1α). Axin acts as a central scaffold protein within this complex, facilitating interactions between GSK-3, CK1α, and β-catenin. Through these interactions, β-catenin undergoes phosphorylation by GSK-3 and CK1α, marking it for subsequent degradation.103
β-catenin is ubiquitinated and degraded by the proteasome system after receiving four phosphates (CK1α adds one phosphate, while GSK-3 adds the three phosphates). CK1α and GSK-3 also phosphorylate Axin and APC in addition to phosphorylating β-catenin in order to promote β-catenin binding and structural stability of the complex.104-106
Wnt binds to the Frizzled receptor (Fz), a seven-pass transmembrane protein, along with its co-receptor LRP5/6. This interaction triggers a series of key events that regulate the Wnt/β-catenin signaling pathway. In other words, Wnt binding causes CK1α and GSK-3 to phosphorylate LRP5/6. This phosphorylation recruits these kinases to the plasma membrane and enhances their binding to LRP5/6.107,108 It should be noted that Dishevelled, activated by Frizzled, is crucial for facilitating the interaction of Axin and GSK-3 with LRP5/6. Cytoplasmic accumulation of β-catenin induces translocation to the nucleus to activate transcription factors of the T-cell factor/lymphoid enhancer factor (TCF/LEF) family (Fig. 3).103
Investigating the impact of the Wnt signaling pathway on facilitating EMT can be approached from three key perspectives: (i) upregulation of EMT-associated transcription factors (such as Zeb and Snail), (ii) disruption of cell-to-cell junctions via preventing β-catenin phosphorylation, and (iii) promotion of stem cell-like characteristics. In triple-negative breast cancer (TNBC), miR-506-3p functions as a tumor suppressor; however, its activity is reduced by circ-0008784, which results in increased activation of the Wnt/β-catenin pathway. Furthermore, circ-0008784 promotes oncogenic properties and upregulates mesenchymal markers.109 Conversely, circ-ITCH, which suppresses miR-214 and miR-17, counteracts TNBC's oncogenic properties by overwhelming the Wnt/β-catenin pathway.110 Recent research has shed light on the pivotal role of CircABCC4 in the progression of breast cancer. Its regulatory role in promoting the activation of NF-κB and Wnt/β-catenin signaling pathways through miR-154-5p inhibition, coupled with the suppression of apoptosis and enhancement of metastatic potential in cancer cells, has been comprehensively elucidated.111 On the other hand, investigations have confirmed an elevated expression pattern of circRNA-069718 in patients with TNBC. CircRNA-069718 knockdown has demonstrated a notable reduction in the levels of key components of the Wnt/β-catenin pathway.112 CircPSMA1, by inhibiting miR-637, is implicated in promoting the proliferation and metastasis of TNBC. This is expected to occur through upregulating β-catenin expression via Akt1, which is a target of miR-637.113 CircRNA seldom yields peptides, but instances like the production of EIF6-224aa from hsa-circ-0060055 (circ-EIF6) demonstrate its possibility. EIF6-224aa enhances MYH9 stability, thereby bolstering the activity of the Wnt/beta-catenin pathway. While the circ-EIF6 oncogenic role in breast cancer is determined, its association with EMT warrants further investigation.114
Interactions between circRNAs and the EMT pathway have been observed in various malignancies, including colorectal cancer. In this context, circPDSS1 and circ-0026628 have been found to promote Wnt/β-catenin-induced EMT by suppressing miR-320c and miR-346, respectively.115,116 Conversely, increasing levels of circ-0026344 have been shown to inhibit EMT by targeting the miR-183-dependent Wnt/β-catenin signaling pathway in colorectal cancer. Additionally, the inhibition of EMT in the case of increasing circ-0026344 by hindering the miR-183-dependent Wnt/β-catenin signaling has been reported in colorectal cancer.117
Similar to the previous signaling pathway, targeted research in this area can also be proposed. For example, in colorectal cancer, the upregulation of circ-0038718 leads to increased expression of Axin2 by inhibiting miR-195-5p. This observation further highlights the intricate connection between circRNAs and the EMT pathway.118 Hsa-circ-000984 and hsa-circ-0007059 have opposite effects on lung cancer cell’s potential to metastasis, as evidenced by their relative inhibitory and acceleratory effects on Wnt/β-catenin-induced EMT, respectively.119,120 These findings, which summarized in Fig. 3, demonstrate that circRNAs critically modulate Wnt/β-catenin signaling to drive EMT, tumor cell invasion, and metastatic spread.
PI3K/AKT mediated regulation of EMT
The initiation of PI3K activity is primarily triggered by ligand binding to receptors such as receptor tyrosine kinases (RTKs) or G-protein-coupled receptors (GPCRs), which serves as the fundamental step for downstream biological processes.121 This cascade entails the conversion of PIP2 (phosphatidylinositol 4,5-bisphosphate) into PIP3 (phosphatidylinositol (3,4,5)-trisphosphate) via PI3K-mediated phosphorylation. This event crucially localizes and activates AKT (protein kinase B) within the plasma membrane.103
Activation of AKT via phosphorylation by phosphoinositide-dependent kinase 1 (PDK1) and mammalian target of rapamycin complex 2 (mTORC2) triggers its full activation, unleashing a cascade of downstream effectors. These include mTORC1, GSK-3, Bad, and FOXO, regulating processes that are implicated in growth, cell survival, homeostasis, and gene expression.122
The primary impact of AKT on the EMT process lies in its kinase function in transcription factor activation like, Snail, Slug, Twist1/2, and Zeb1/2.123 Furthermore, AKT with GSK-3 phosphorylation ensures stability and cytoplasmic accumulation of β-catenin.124 On the other hand, AKT also serves a central role in modulating key signaling pathways involved in EMT, including the Wnt/β-catenin and TGF-β pathways.
Considering the oncogenic nature of the PI3K/AKT signaling pathway and the positive link to EMT, exploring how the circRNA/miRNA axis affects EMT via this pathway is promising. Reduced levels of hsa-circ-001569 in breast cancer result in diminished malignancy-associated traits, including decreased activity of the PI3K/AKT pathway, lower expression levels of EMT markers at the molecular level, and reduced proliferation. Consequently, it presents a promising therapeutic target for enhancing prognosis in breast cancer cases.125
In breast cancer, circSEMA4B functions oppositely by impeding AKT phosphorylation, thereby exerting inhibitory effects on the EMT process.126 Research has elucidated that miR-338-3p exerts a suppressive function on EMT by targeting Zeb2 in breast cancer.127 In parallel, circKIF4A is a suppressor of metastasis in TNBC by inhibiting miR-375.128 Interestingly, PDK1 was mentioned as a target of miR-375 in pancreatic cancer.129 However, while the inhibitory effect of circ-0000326 on miR-338 has been established in bladder cancer, this relationship remains unexplored in breast cancer, highlighting a notable gap in current understanding.130 Furthermore, research exploring the relationship between hsa-circ-0003288 and N-cadherin expression levels in hepatocellular carcinoma has demonstrated a supportive role in EMT (Fig. 4).131 Overall, these studies reveal that circRNAs critically orchestrate PI3K/AKT pathway activity, either by sponging miRNAs or modulating key kinases, to regulate EMT and metastatic behavior. Table 2 presents a summary of additional relevant studies within this field.
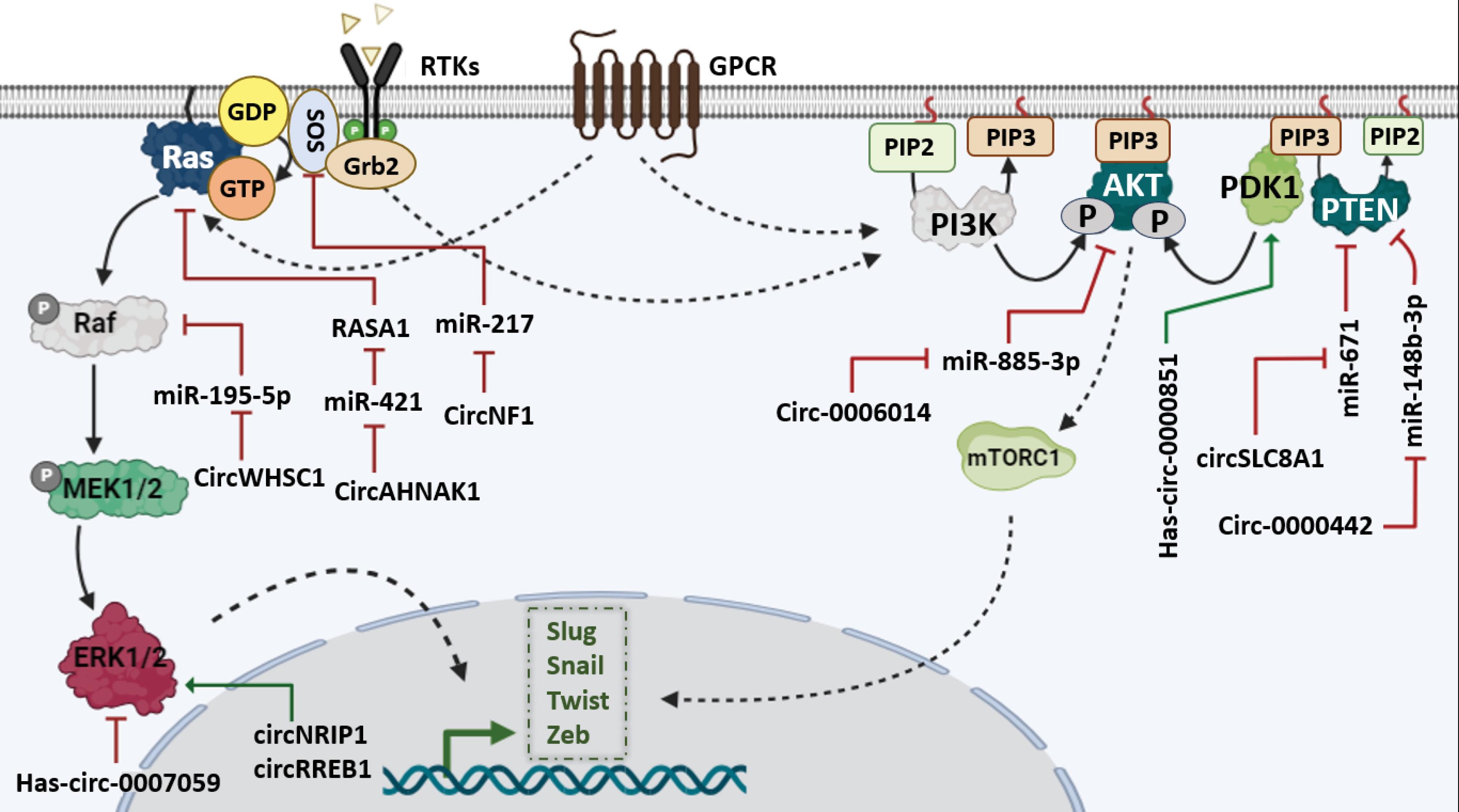
Fig. 4.
Schematic picture of the interaction of circRNAs with MAPK-MEK and PI3K/AKT signaling pathways.
.
Schematic picture of the interaction of circRNAs with MAPK-MEK and PI3K/AKT signaling pathways.
Table 2.
summary of circRNAs implication in the PI3K–EMT network
|
circRNA
|
Role
|
Cancer
|
Target miRNA
|
Pathway-related miRNA Target
|
Ref.
|
| circSLC8A1 |
Anti-tumor |
Breast cancer |
miR-671 |
PTEN |
132
|
| circ-0000442 |
Anti-tumor |
Breast cancer |
miR-148b-3p |
PTEN |
133
|
| circ-0006014 |
pro-tumor |
Breast cancer |
miR-885-3p |
NTRK2/PIK3 |
134
|
| hsa-circ-0000851 |
pro-tumor |
triple-negative breast cancer |
miR-1183 |
PDK1/p-AKT |
135
|
| circWAC |
pro-tumor |
triple-negative breast cancer |
miR-142 |
WWP1/PI3K |
136
|
| circ-100395 |
Anti-tumor |
gastric cancer |
miR-142-3p |
PTEN |
137
|
MAPK-MEK mediated regulation of EMT
One of the most important members of the cellular signaling network is the classical MAP kinase pathway, which is also known as the Ras/Raf/MEK/ERK cascade. It is involved in biological processes including apoptosis, autophagy, differentiation, growth, and metastasis. Stimulation of receptors, mostly GPCRs and RTKs, leads to the activation of RAS, a monomeric G-protein, which is mediated by GRB2 and SOS. Active Ras activates a serine-threonine kinase called Raf.138 In turn, Raf phosphorylates and activates a dual-specificity kinase called MAPK/ERK kinase or mitogen-activated protein kinase (MEK). By phosphorylating ERK, MEK enables the phosphorylation of a wide range of cytoplasmic/nuclear mediators by ERK.139
Induction of transcription factors related to EMT, regulation of cytoskeleton and cell-to-cell connection, increase of cell resistance to apoptosis, and regulation of tumor environment to facilitate metastasis (for example, by increasing MMPs) are among the most important effects of the MAPK pathway on progression of EMT in breast cancer. Besides this, the interaction with other pathways facilitating EMT, such as TGF-β, Wnt/β-catenin, and Notch, shows another dimension of the effect of the map kinase pathway on EMT.140-142
Research into the connection between circRNAs and EMT in breast cancer has highlighted the significant roles of circAHNAK1 and circRREB1.143 CircRREB1 promotes breast cancer progression by enhancing tumor cell migration and proliferation while inhibiting apoptosis, with activation of ERK1/2 identified as a key underlying mechanism. Conversely, circAHNAK1 is downregulated in triple-negative breast cancer, which abolishes its tumor-suppressive effect. By failing to inhibit miR-421, reduced circAHNAK1 expression leads to decreased RASA1 activity, thereby enhancing Ras-mediated signaling and promoting cancerous traits such as proliferation and metastasis.144,145
Other circRNAs implicated in the MAPK-EMT network are summarized in Table 3. Collectively, these findings underscore the crucial regulatory roles of circRNAs in breast cancer metastasis and highlight the circRNA-MAPK axis as a potential biomarker for assessing tumor progression.
Table 3.
summary of circRNAs implication in the MAPK-EMT network
|
circRNA
|
Role
|
Cancer
|
Target miRNA
|
Pathway-related miRNA Target
|
Ref.
|
| circWHSC1 |
Pro-tumor |
breast cancer |
miR-195-5p |
FASN/AMPK |
146
|
| circ-0042881 (circNF1) |
Pro-tumor |
breast cancer |
miR-217 |
SOS1 |
147
|
| circAHNAK1 |
Anti-tumor |
triple-negative breast cancer |
miR-421 |
RASA1/Ras |
145
|
| hsa-circ-0007059 |
Anti-tumor |
lung cancer |
miR-378 |
Wnt/β-catenin and ERK1/2 |
120
|
| circ-ZKSCAN1 |
Pro-tumor |
non-small cell lung cancer |
miR-330-5p |
FAM83A |
148
|
| circNRIP1 |
Pro-tumor |
gastric cancer |
miR-149-5p |
AKT1 |
149
|
| Pro-tumor |
cervical cancer |
miR-629-3p |
ERK1/2 |
150
|
| circ-0087558 |
Pro-tumor |
breast cancer |
miR-643 |
MAP2K6 |
151
|
Other pathways-mediated regulation of EMT
The binding of STAT3 to an active receptor leads to STAT3 phosphorylation and dimerization. After then, dimerized STAT3 is translocated to the nucleus, where it attaches to STAT-binding elements (SBEs) to start the transcription of different genes.152 Studies have found that elevated levels of circNOLC1 in breast cancer are correlated with decreased E-cadherin and increased vimentin expression. The mechanism by which circNOLC1 enhances the oncogenic characteristics and metastatic potential of breast cancer involves acting as a sponge for miR-365a-3p, thereby sustaining STAT3 activity.153 Increased expression of circSEPT9 in TNBC is also associated with poor prognosis. The mechanism of circSEPT9 in increasing the occurrence of oncogenic properties, including metastasis, is by inhibiting miR-637 in order to sustain the activity of the Leukemia Inhibitory Factor (LIF)/STAT3 axis.154 Other malignancies have reported an increase in STAT3 activity associated with circ-0068871, acting as a sponge for miR-124; ciRS-7, acting as a sponge for miR-7; circFCHO2, acting as a sponge for miR-194-5p; and hsa-circ-0000043, acting as a sponge for miR-4492.155-159
The notch signaling pathway initiates by binding the notch receptor (Notch-1, -2, -3, and -4) in a cell to the ligand in the adjacent cell. This connection triggers proteolytic reactions that release the intracellular part of the notch receptor (NICD). In turn, NICD transfers to the nucleus and eventually increases the expression of the notch target genes.103 Although studies have not presented a clear connection between the Notch signaling pathway and circRNA in mediating EMT in breast cancer, recent studies have shed light on the role of hsa-circRNA-102051, circKIF4A, circAPLP2, and circPDK1 in the activation of the Notch pathway by inhibiting miR-203a, miR-375/1231, miR-101-3p, and miR-377-3p, respectively. Conversely, circ-0000390 inhibits the notch as a tumor suppressor. However, these findings require further validation through additional investigations in the field of breast cancer.160-164
The Hedgehog signaling pathway starts with connecting secretory ligands (i.e., Hedgehog proteins) to a membrane receptor called a Patched. This connection removes the Patched inhibition effect from the Smoothened. This signaling cascade eventually leads to the activation of transcription factors of the Glioma-associated oncogene (Gli) family, which can increase the molecular level of transcription factors associated with EMT.103 CircZNF609 in hepatocellular carcinoma promotes Hedgehog pathway activation by suppressing miR-15a-5p/15b-5p, while Circ-0000034 in retinoblastoma enhances this pathway through upregulation of ADAM19 via sponging miR-361-3p.165,166 Although a substantial portion of circRNA interactions with other signaling pathways remains uncharacterized, the limited evidence available highlights the urgent need to direct research efforts towards this domain.
Circular RNAs in the regulation of late-stage; post-EMT events
Following the occurrence of EMT, events such as invasion, angiogenesis, immune evasion, and colonization at distant sites play a pivotal role in the success of metastasis. In this context, circRNAs act as critical regulators orchestrating these post-EMT processes.16
The ECM, composed of both fibrous and non-fibrous components, acts as a protective barrier within tissues, preventing the spread and invasion of cancer cells. ECM remodeling is essential for enhancing the ability of cancer cells to infiltrate the circulatory system and evade immune surveillance during metastasis, as it facilitates the transmission of oncogenic signals. A prominent example is circACAP2, an oncogenic circRNA that is upregulated in breast cancer and promotes the expression of the COL5A1 gene by inhibiting miR-29a-3p and miR-29b-3p.167 Qu et al demonstrated that the upregulated activity of circRNA-CER causes the high expression of MMP13 through a mechanism dependent on the inhibition of miR-136, consequently accelerating ECM remodeling.168 Invasion involves specialized actin-rich protrusions known as invadopodium, which cancer cells utilize to degrade the ECM and infiltrate surrounding tissues. A study by Du WW et al. demonstrated that circSKA3 promotes invadopodium formation by complexing with Tks5 and integrin β1.169 It is noteworthy that alongside circRNAs that activate invasiveness, such as hsa_circ_0052112 and circ_0103552, some antitumor circRNAs like circASS1 attenuate the invasive phenotype.170-172
The interaction of circRNAs with the immune system in order to facilitate the escape of cancer cells from the immune system or to inhibit this system for the development of cancer is a very noteworthy area in cancer studies. In breast cancer research, the investigation into the role of circ-002172 serves as a striking illustration of circRNA’s interaction with the immune system. This upregulated circRNA not only suppresses the infiltration of cytotoxic T cells but also counteracts the inhibitory effect of miR-296-5p, consequently enhancing the expression of CXCL12. While CXCL12 impacts both the innate and adaptive components of the immune system, its precise role in cancer development remains uncertain.173 Another example in this regard is circWWC3, which raises the possibility of breast cancer metastasizing by raising the molecular level of interleukin-4.174 In breast cancer, upregulated hsa_circ_0067842 promotes the stabilization of CMTM6, thereby impeding the ubiquitination-mediated degradation of PD-L1. The subsequent binding of PD-L1 to PD-1 on T cells attenuates antitumor immunity by driving T cell exhaustion.175 Another group of circRNAs, including circ_0001598, circ_0000512, circATAD2, and circGSK3β, has been shown to elevate PD-L1 expression levels, thereby suppressing CD8⁺ T cell-mediated antitumor immune surveillance in breast cancer. However, current studies do not specifically attribute this immune evasion to the metastatic process alone.176-179
Angiogenesis is essential in the metastatic process, providing cancer cells with the necessary resources to survive.180 According to reports, hsa-circRNA-002178 in breast cancer increases angiogenesis and worsens prognosis by suppressing the inhibitory effect on COL1A1 through a sponge-like function for miR-328-3p.181 On the other hand, exosomal lncRNASNHG12 has been shown to promote angiogenesis in breast cancer through the PBRM1/MMP10 axis.182
CircRNAs play a multifaceted role in creating favorable conditions for metastatic cell colonization. Their involvement in regulating various processes such as angiogenesis, ECM remodeling, EMT, and modulation of immune response in the tumor microenvironment (TME) collectively promotes colonization potential in an indirect manner.183,184
It is important to note that although the role of circRNAs in individual post-EMT processes such as invasion, immune evasion, ECM remodeling, and angiogenesis is well-documented, these mechanisms have largely been studied in isolation. A more integrated investigation that considers their coordinated contribution within the full metastatic cascade may yield deeper insights into the functional relevance of circRNAs in breast cancer progression.
Translational and clinical relevance of circular RNA in breast cancer
The remarkable stability and abundance of circRNAs in physiological fluids such as blood and urine have highlighted their clinical significance as candidates for liquid biopsy-based diagnostics. Aberrant expression patterns of circRNAs in breast cancer tissues, along with their tissue-specific characteristics, underscore their potential as reliable biomarkers for diagnosis and prognosis, therapeutic targets, and mediators of treatment resistance.185 In addition, the differentiation of subtypes of breast cancer is possible based on the expression of circRNAs. For instance, circ-0044234 expression is notably lower in TNBC compared to other breast cancer subtypes, suggesting its utility in subtype differentiation.186
Although no circRNA-based panel has yet been implemented in routine clinical practice for breast cancer diagnosis, accumulating evidence underscores their substantial diagnostic potential. In support of this, Libin et al showed that in 136 breast cancer patients, hsa_circ_0006969 achieved AUCs of 0.889 in tumor tissue and 0.823 in blood; 0.672 and 0.675 for grades 1 and 3; 0.648 in ER-positive cases; 0.687–0.757 across TNM stages I–III (0.751 for stages I–II); and outperformed CA15-3 in detecting metastasis (0.742 vs. 0.712).187 Similarly, the ROC analysis of hsa_circ_0000615 demonstrated an AUC of 0.904 (95% CI: 0.863–0.944), with a sensitivity of 76.8% and specificity of 88.4% in distinguishing non-metastatic breast cancer cases.188 Moreover, hsa_circ_0001785 in plasma showed a diagnostic AUC of 0.784.189 Consistently, several studies have identified circRNAs as promising diagnostic and prognostic biomarkers for TNBC. For instance, hsa_circ_0072309 is downregulated in TNBC and discriminated patients from healthy controls with AUCs of 0.78 (FFPE) and 0.81 (frozen), and its activity appears linked to interactions with RNA-binding proteins affecting PTEN and RUNX1 signaling.190 circZCCHC2 and circCAPG are upregulated in breast tumors including TNBC, with diagnostic AUCs of 0.787 and 0.8723, respectively; circCAPG is furthermore associated with poorer overall survival.191 From a prognostic perspective, elevated circWHSC1 (AUC 0.968) correlates with shorter overall survival in TNBC, and serum circPRMT5 (AUC 0.9323) is associated with metastasis and treatment resistance, highlighting their potential for noninvasive monitoring.191 These candidates warrant validation in larger cohorts and mechanistic studies.
Several circRNA profiles identified in breast cancer have been proposed as prognostic markers in prior research. Elevated levels of oncogenic circRNAs like circEPSTI1 and circKIF4A are associated with a worse prognosis, whereas increased expression of tumor-suppressive circRNAs such as circ-VRK1 correlates with improved outcomes for breast cancer patients.192-194 Specifically, patients with high circEPSTI1 expression showed significantly shorter mean disease-free survival and overall survival compared to those with low expression levels.192 It should be noted that, according to ClinicalTrials.gov, an observational study (NCT05771337) is currently recruiting to evaluate the diagnostic and prognostic value of hsa_circ_0001785 and hsa_circ_100219 in serum samples from breast cancer patients, with estimated completion in 2025.195
On the other hand, circRNAs contribute pointedly to drug resistance and restrict treatment options in breast cancer by regulating multiple cellular processes such as autophagy, apoptosis, cell cycle progression, DNA repair, and drug efflux. This sustains the survival of cancer cells, contributing to increased mortality rates in breast cancer patients. Research in this field has elucidated the involvement of specific circRNAs in drug resistance mechanisms: circ-BGN in resistance to trastuzumab, circ-MMP11 in resistance to lapatinib, hsa-circ-0092276 in promoting autophagy and resistance to doxorubicin, circ-CDR1as in enhancing resistance to chemotherapy, and circPVT1 in augmenting drug resistance specifically in the ER + subtype. Consequently, circRNA expression profiling in breast tumors holds significant promise for predicting drug resistance patterns and informing the selection of tailored therapeutic strategies. Such precision-guided approaches may not only improve treatment outcomes but also accelerate the integration of personalized medicine into breast cancer care.196-200
As previously mentioned, circRNAs regulate breast cancer progression primarily by functioning as competing endogenous RNAs, which sponge tumor-suppressor or oncogenic miRNAs. This, in turn, modulates key signaling cascades such as PI3K/AKT, Wnt/β-catenin, and MAPK/ERK. In this regard, oncogenic Hsa_circ_001569 and CirCHIPK3 promote proliferation and metastasis via PI3K/AKT activation, while circRNA-SFMBT2 and Circ-RNF111 drive therapy resistance by sequestering miRNAs that would otherwise sensitize tumors to chemotherapy. Targeted inhibition of these circRNAs in preclinical models has restored drug sensitivity and suppressed tumor characteristics, underscoring their potential as both biomarkers of resistance and actionable targets for precision therapies in breast cancer.201-204 Another example in this area is the silencing of circAGFG1 using antisense oligonucleotides, which significantly inhibited proliferation and EMT in vitro, while also restoring sensitivity to chemotherapy.205 Despite advances in engineering exosomes for targeted delivery of antitumor compounds such as miRNAs, this field remains largely unexplored for tumor-suppressive circRNAs.
Despite growing interest in the role of circRNAs in cancer-related EMT, much of the available evidence remains confined to in vitro studies using cancer cell lines or limited tissue-level analyses. In many cases, investigations have focused solely on altered circRNA expression and their impact on migration or proliferation, without extending these observations to in vivo models or patient-derived data. While notable exceptions exist, such as circANKS1B,206 which has been functionally validated in tumor xenograft models and shown to promote in vivo tumorigenesis, most circRNAs proposed to regulate EMT lack such mechanistic or translational validation. As a result of these preclinical gaps, the clinical translation of circRNA biomarkers remains delayed by the absence of standardized and integrated analytical frameworks. Without precise data‐quality metrics, targeted enrichment strategies and ultra-sensitive platforms (e.g., ddPCR), unified SOPs for isolation and quantification, and consensus‐driven multicenter validation, reproducibility suffers and diagnostic adoption is hindered.
Interpretation of findings: functional discrepancy
CircRNAs frequently behave paradoxically: the same circular transcript can act as a tumor promoter in one context and as a suppressor in another. Early studies largely reduced circRNAs to “miRNA sponges,” but mounting evidence indicates that protein interactions, translation of circRNA-derived peptides, and other post-transcriptional mechanisms often play equal or greater roles. Take the TGFβ/SMAD axis as an example. In breast cancer models, circANKS1B and circ-0000043 have been reported to drive EMT by sequestering miR-148a-3p/miR-152-3p and miR-136, thereby amplifying Smad-dependent mesenchymal programs.94,96 Yet other tumor types tell a different story: circPVT1 upregulates Smad3 and promotes EMT in gastric cancer,97 whereas circ-OXCT1 suppresses EMT through Smad4 inhibition.98 Such cross-tissue contrasts are likely rooted in differences in epigenetic state and baseline pathway activity (for instance, tissue-specific TGFβ levels) which make straightforward extrapolation hazardous. For meaningful interpretation of circRNA function in breast cancer, we therefore need experiments conducted in situ and in models that preserve tumor subtype identity and microenvironmental context. Only then can we distinguish context-specific biology from artifacts of simplified systems.
A comparable functional divergence emerges within the Wnt/β-catenin pathway. For example, circ-0008784 and circABCC4 have been reported to promote EMT by sequestering miR-506-3p and miR-154-5p, respectively; whereas circ-ITCH appears to counteract EMT through inhibition of miR-214 and miR-17.109-111 Importantly, most evidence supporting circ-ITCH comes from in vitro systems. Without in vivo corroboration, its perceived effect size may be inflated because complex TME interactions are not captured. Studies in non-breast malignancies echo this heterogeneity. CircPDSS1 and circ-0026628 drive Wnt-dependent EMT in colorectal and lung cancers, while circ-0026344 exerts an inhibitory influence.115-117 Assertions that these opposing roles are evolutionarily conserved remain premature in the absence of phylogenetic analyses and may reflect publication bias. Adding another layer of complexity, some circRNAs encode peptides (for instance, EIF6-224aa from circ-EIF6) which can alter regulatory outcomes depending on isoform abundance and subcellular targeting.114 Because bona fide translation of circRNAs appears infrequent under physiological conditions, rigorous biochemical and functional validation is essential to discriminate genuine biology from experimental artifact.
Parallel contradictions appear in PI3K/AKT and MAPK/MEK signaling. For example, circ-0006014 and hsa-circ-0000851 have been reported to activate PI3K/AKT and thereby promote EMT.134,135 By contrast, circSEMA4B reduces AKT phosphorylation and suppresses the same program.126 Crucially, most of these claims rely on knockdown or overexpression systems. Without quantification of endogenous circRNA abundance, the physiological relevance of such perturbations remains ambiguous.
The picture is further muddled by indirect inferences. CircKIF4A in TNBC has been proposed to restrain PDK1 through miR-375 inhibition,161 yet this link has not been validated in breast tumors. Such extrapolations risk creating a domino effect of untested assumptions and mask potential pathway crosstalk. To move beyond speculation, we need loss- and gain-of-function experiments that precisely target backsplice junctions, for example, CRISPR-based editing.
In MAPK/MEK signaling similar mixed signals emerge. CircWHSC1 appears to enhance Raf1-dependent EMT via miR-195-5p sponging,146 whereas circ-ZKSCAN1 and circNRIP1 act inhibitory in other malignancies.148,150 Do circRNAs operate as finely tunable rheostats, subtly modulating pathway flux? Or do they behave as binary switches that flip cellular states? Framing the question this way is misleadingly simple. A more realistic model places circRNAs inside stochastic, noisy regulatory networks where genetic variation and environmental context shape net output. In practice, resolving these conflicts will require three things: accurate, absolute quantification of endogenous circRNAs; CRISPR-based causal tests; and integrative network modeling that accommodates stochastic behavior and context dependency.
The absence of standardized benchmarks for circRNA-miRNA interactions (for example, agreed-upon binding-affinity or free-energy cutoffs) undermines reproducibility across studies. In silico predictions frequently diverge from biochemical assays, producing spurious target lists. Widely used algorithms (e.g., miRanda, TargetScan) have a well-documented tendency to overcall interactions when used in isolation.207 To avoid false positives, computational screens must be coupled with orthogonal experimental evidence. Integrating transcriptome-wide CLIP datasets, AGO-pull-downs, or direct reporter assays into a unified validation pipeline will greatly improve confidence in functional assignments.
The functional ambiguity of circRNAs extends well beyond EMT. Some circular RNAs, such as circACAP2 and circRNA-CER, drive ECM remodeling and invasion by upregulating COL5A1 and MMP13.167,168 Others, for example circASS1, act in opposition and blunt invasive behavior.172 These opposing effects indicate that circRNAs can exert bidirectional control over metastatic fitness. Critically, investigations that focus narrowly on ECM remodeling frequently overlook immune interactions. CircRNAs may reshape the tumor stroma indirectly via cancer-associated fibroblasts, or by altering chemokine and checkpoint axes that modulate immune surveillance. For example, circ-002172 and hsa_circ_0067842 have been linked to T-cell exhaustion through CXCL12/PD-L1 signaling.173,175 However, whether these immune effects are specific to metastatic niches or also operate in primary tumors is unresolved.
A major translational limitation is model choice. Many immune studies use immunocompromised hosts, which cannot recapitulate human antitumor immunity. Humanized mouse models and complementary ex vivo systems, are therefore essential to validate immune-related claims.
Angiogenesis adds another layer of uncertainty. Reports implicate hsa-circRNA-002178 in neovascularization, yet direct, subtype-stratified comparisons are lacking.181 It remains unclear if pro-angiogenic circRNA activity is universal across breast cancer subtypes or enriched in subsets such as TNBC.
Finally, exosomal circRNAs are an attractive mechanism for intercellular signaling, but their functional role is under-validated. Demonstrating bona fide transfer and biological effect requires rigorous in vivo tracing and quantitative uptake assays. In short, resolving these contradictions demands integrated, physiologically relevant models that couple stromal, immune, and vascular readouts with rigorous in vivo validation.
Taken together, these context-dependent and often contradictory effects underscore a core challenge in circRNA research. Many studies rely heavily on gain- and loss-of-function experiments in conventional cell lines. Such approaches can artificially amplify circRNA functions due to overexpression, while ignoring the natural stoichiometry and compensatory mechanisms present in vivo. As a result, the generalizability of these findings is limited. Relying on simplified models further constrains progress. To address this, more physiologically relevant systems are essential. Organoids, for example, preserve tissue architecture and cellular diversity. Single-cell RNA sequencing can capture heterogeneity across cell types and reveal microenvironmental influences. Integrating these advanced platforms will allow a more accurate and nuanced understanding of circRNA functions in cancer biology.
Conclusion and Future Perspectives
CircRNAs regulate EMT in breast cancer through several distinct mechanisms. These include functioning as competing endogenous RNAs (ceRNAs) to sponge miRNAs, interacting with RNA-binding proteins, modulating transcription, influencing alternative splicing, and affecting post-transcriptional gene regulation. Collectively, these categories illustrate the diverse ways in which circRNAs can impact EMT and contribute to breast cancer progression.
In parallel, circRNAs have also emerged as pivotal regulators of breast cancer metastasis-related signaling pathways. Comprehensive profiling of circRNAs in metastatic breast cancer patients is a critical first step towards developing circRNA-directed therapies. Promising strategies include silencing oncogenic circRNAs and delivering tumor-suppressive circRNAs via bioengineered exosomes as adjuncts to existing treatments. However, several challenges must be addressed before clinical translation can occur. These include incomplete functional characterization of many circRNAs, a lack of standardized analytical protocols, hurdles in achieving targeted tissue delivery, the inherent complexity of circRNA-regulatory networks, limited clinical validation, and regulatory and manufacturing barriers.
Looking ahead, integrating circRNA research with artificial intelligence and high-throughput computational platforms promises to uncover novel regulatory circuits through machine-learning–driven network analysis. Such integration can enhance metastasis-prediction models by incorporating circRNA signatures into prognostic algorithms and enable earlier detection of aggressive tumor subtypes via multi-omic biomarker panels. Establishing robust, interoperable circRNA databases will be instrumental in advancing these efforts.
Ultimately, circRNAs represent a cutting-edge frontier in precision oncology, offering new avenues to overcome drug resistance, inhibit metastatic spread, and improve clinical outcomes for patients battling advanced breast cancer.
Review Highlights
What is the current knowledge?
-
Circular RNAs act as miRNA sponges and regulate gene expression in cancers.
-
EMT drives breast cancer metastasis via TGF-β, Wnt/β-catenin, PI3K/AKT, and MAPK pathways.
-
circRNA–miRNA axes modulate signaling components controlling EMT and metastatic traits.
-
circRNAs influence post-EMT processes: ECM remodeling, invasion, angiogenesis, and immune evasion.
-
Many circRNA–EMT interactions are characterized in other cancers but underexplored in breast cancer.
What is new here?
-
This review synthesizes circRNA-mediated regulation of EMT specifically in breast cancer metastasis.
-
We highlight circANKS1B, circ-0000043 and others that promote EMT via miRNA/Smad pathways.
-
Evidence shows circRNAs both activate and inhibit Wnt, PI3K/AKT, and MAPK signaling affecting EMT.
-
We emphasize circRNAs' roles in post-EMT events and potential therapeutic targeting opportunities.
-
We propose targeted research directions to validate circRNA-based biomarkers and anti-metastatic strategies.
Competing Interests
The authors declare no conflict of interest.
Consent for Publication
Not applicable.
Data Availability Statement
Not applicable.
Declaration of AI-assisted Tools in the Writing Procedure
The authors declare that ChatGPT (OpenAI, San Francisco, CA, USA) was used solely for language editing and improving the readability of the manuscript. The tool was not involved in the generation of scientific content, data analysis, or interpretation of results.
Ethical Approval
Not applicable.
References
- Jesinger RA. Breast anatomy for the interventionalist. Tech Vasc Interv Radiol 2014; 17:3-9. doi: 10.1053/j.tvir.2013.12.002 [Crossref] [ Google Scholar]
- Khan YS, Fakoya AO, Sajjad H. Anatomy, Thorax: Mammary Gland. StatPearls. Treasure Island (FL): StatPearls Publishing Copyright © 2026, StatPearls Publishing LLC.; 2026.
- Malhotra GK, Zhao X, Band H, Band V. Histological, molecular and functional subtypes of breast cancers. Cancer Biol Ther 2010; 10:955-60. doi: 10.4161/cbt.10.10.13879 [Crossref] [ Google Scholar]
-
Christgen M, Cserni G, Floris G, Marchio C, Djerroudi L, Kreipe H, et al. Lobular Breast Cancer: Histomorphology and Different Concepts of a Special Spectrum of Tumors. Cancers (Basel) 2021; 13. doi: 10.3390/cancers13153695.
- Makki J. Diversity of Breast Carcinoma: Histological Subtypes and Clinical Relevance. Clin Med Insights Pathol 2015; 8:23-31. doi: 10.4137/CPath.S31563 [Crossref] [ Google Scholar]
- Alluri P, Newman LA. Basal-like and triple-negative breast cancers: searching for positives among many negatives. Surg Oncol Clin N Am 2014; 23:567-77. doi: 10.1016/j.soc.2014.03.003 [Crossref] [ Google Scholar]
- Harbeck N, Penault-Llorca F, Cortes J, Gnant M, Houssami N, Poortmans P. Breast cancer. Nat Rev Dis Primers 2019; 5:66. doi: 10.1038/s41572-019-0111-2 [Crossref] [ Google Scholar]
-
Dogan I, Yirgin IK, Ozkurt S, Ibıs K, Vatansever S, Saip P, et al. Outcomes and prognostic factors in patients with HER2-positive metastatic breast cancer with brain metastasis. Journal of Clinical Oncology. 40: e14002–e. doi: 10.1200/JCO.2022.40.16_suppl.e14002.
- Orrù S, Pascariello E, Sotgiu G, Piras D, Saderi L, Muroni MR, et al. Prognostic Role of Androgen Receptor Expression in HER2 + Breast Carcinoma Subtypes. Biomedicines 2022; 10. doi: 10.3390/biomedicines10010164.
-
Bennett C, Carroll C, Wright C, Awad B, Park JM, Farmer M, et al. Breast Cancer Genomics: Primary and Most Common Metastases. Cancers (Basel) 2022; 14. doi: 10.3390/cancers14133046.
-
Fontanella C, Gade S, Von Minckwitz G, Lederer B, Blohmer JU, Costa SD, et al. Prognostic score for Luminal A-like breast cancer patients. Journal of Clinical Oncology. 32: 525–. doi: 10.1200/jco.2014.32.15_suppl.525.
- Rizzo A, Cusmai A, Massafra R, Bove S, Comes MC, Fanizzi A. Systemic Treatments for Metastatic Human Epidermal Growth Factor Receptor 2-Positive Breast Cancer: Old Certainties and New Frontiers. Cancer Control 2022; 29:10732748221106267. doi: 10.1177/10732748221106267 [Crossref] [ Google Scholar]
- Chen W, Hoffmann AD, Liu H, Liu X. Organotropism: new insights into molecular mechanisms of breast cancer metastasis. NPJ Precis Oncol 2018; 2:4. doi: 10.1038/s41698-018-0047-0 [Crossref] [ Google Scholar]
- Cohen SY, Stoll CR, Anandarajah A, Doering M, Colditz GA. Modifiable risk factors in women at high risk of breast cancer: a systematic review. Breast Cancer Res 2023; 25:45. doi: 10.1186/s13058-023-01636-1 [Crossref] [ Google Scholar]
- Sun YS, Zhao Z, Yang ZN, Xu F, Lu HJ, Zhu ZY. Risk Factors and Preventions of Breast Cancer. Int J Biol Sci 2017; 13:1387-97. doi: 10.7150/ijbs.21635 [Crossref] [ Google Scholar]
- Molavand M, Montazer M, Ghareghomi S, Yousefi B, Majidinia M. The involvement of major signaling pathways in metastasis of colorectal cancer and the therapeutic application of their inhibitors. Expert Rev Anticancer Ther 2025; 25:915-38. doi: 10.1080/14737140.2025.2519860 [Crossref] [ Google Scholar]
- Liu Z, Chen J, Ren Y, Liu S, Ba Y, Zuo A. Multi-stage mechanisms of tumor metastasis and therapeutic strategies. Signal Transduct Target Ther 2024; 9:270. doi: 10.1038/s41392-024-01955-5 [Crossref] [ Google Scholar]
- Chen Y, Lu L, Feng B, Han S, Cui S, Chu X. Non-coding RNAs as emerging regulators of epithelial to mesenchymal transition in non-small cell lung cancer. Oncotarget 2017; 8:36787-99. doi: 10.18632/oncotarget.16375 [Crossref] [ Google Scholar]
- Misir S, Wu N, Yang BB. Specific expression and functions of circular RNAs. Cell Death Differ 2022; 29:481-91. doi: 10.1038/s41418-022-00948-7 [Crossref] [ Google Scholar]
- Molavand M, Ebrahimnezhade N, Kiani A, Yousefi B, Nazari A, Majidinia M. Regulation of autophagy by non-coding RNAs in human glioblastoma. Med Oncol 2024; 41:260. doi: 10.1007/s12032-024-02513-3 [Crossref] [ Google Scholar]
-
Brodowska A, Grabowska M, Bittel K, Ciećwież S, Brodowski J, Szczuko M, et al. Estrogen and Progesterone Receptor Immunoexpression in Fallopian Tubes among Postmenopausal Women Based on Time since the Last Menstrual Period. Int J Environ Res Public Health 2021; 18. doi: 10.3390/ijerph18179195.
- Lange CA, Yee D. Progesterone and Breast Cancer. Women's Health 2008; 4:151-62. doi: 10.2217/17455057.4.2.151 [Crossref] [ Google Scholar]
- Aitken SJ, Thomas JS, Langdon SP, Harrison DJ, Faratian D. Quantitative analysis of changes in ER, PR and HER2 expression in primary breast cancer and paired nodal metastases. Ann Oncol 2010; 21:1254-61. doi: 10.1093/annonc/mdp427 [Crossref] [ Google Scholar]
-
Chimento A, De Luca A, Avena P, De Amicis F, Casaburi I, Sirianni R, et al. Estrogen Receptors-Mediated Apoptosis in Hormone-Dependent Cancers. Int J Mol Sci 2022; 23. doi: 10.3390/ijms23031242.
- Lappano R, Malaguarnera R, Belfiore A, Maggiolini M. Recent advances on the stimulatory effects of metals in breast cancer. Mol Cell Endocrinol 2017; 457:49-56. doi: 10.1016/j.mce.2016.10.017 [Crossref] [ Google Scholar]
- Tarhonska K, Lesicka M, Janasik B, Roszak J, Reszka E, Braun M. Cadmium and breast cancer - Current state and research gaps in the underlying mechanisms. Toxicol Lett 2022; 361:29-42. doi: 10.1016/j.toxlet.2022.03.003 [Crossref] [ Google Scholar]
- Hiller TWR, O'Sullivan DE, Brenner DR, Peters CE, King WD. Solar Ultraviolet Radiation and Breast Cancer Risk: A Systematic Review and Meta-Analysis. Environ Health Perspect 2020; 128:16002. doi: 10.1289/ehp4861 [Crossref] [ Google Scholar]
- Helm JS, Rudel RA. Adverse outcome pathways for ionizing radiation and breast cancer involve direct and indirect DNA damage, oxidative stress, inflammation, genomic instability, and interaction with hormonal regulation of the breast. Arch Toxicol 2020; 94:1511-49. doi: 10.1007/s00204-020-02752-z [Crossref] [ Google Scholar]
- Miao Y, Rong M, Li M, He H, Zhang L, Zhang S. Serum concentrations of organochlorine pesticides, biomarkers of oxidative stress, and risk of breast cancer. Environ Pollut 2021; 286:117386. doi: 10.1016/j.envpol.2021.117386 [Crossref] [ Google Scholar]
- Rizzolo P, Silvestri V, Falchetti M, Ottini L. Inherited and acquired alterations in development of breast cancer. Appl Clin Genet 2011; 4:145-58. doi: 10.2147/tacg.S13226 [Crossref] [ Google Scholar]
- Easton DF, Pharoah PD, Antoniou AC, Tischkowitz M, Tavtigian SV, Nathanson KL. Gene-panel sequencing and the prediction of breast-cancer risk. N Engl J Med 2015; 372:2243-57. doi: 10.1056/NEJMsr1501341 [Crossref] [ Google Scholar]
- Abu-Helalah M, Azab B, Mubaidin R, Ali D, Jafar H, Alshraideh H. BRCA1 and BRCA2 genes mutations among high risk breast cancer patients in Jordan. Sci Rep 2020; 10:17573. doi: 10.1038/s41598-020-74250-2 [Crossref] [ Google Scholar]
- Sheikh A, Hussain SA, Ghori Q, Naeem N, Fazil A, Giri S. The spectrum of genetic mutations in breast cancer. Asian Pac J Cancer Prev 2015; 16:2177-85. doi: 10.7314/apjcp.2015.16.6.2177 [Crossref] [ Google Scholar]
- Weigelt B, Peterse JL, van 't Veer LJ. Breast cancer metastasis: markers and models. Nat Rev Cancer 2005; 5:591-602. doi: 10.1038/nrc1670 [Crossref] [ Google Scholar]
-
Park M, Kim D, Ko S, Kim A, Mo K, Yoon H. Breast Cancer Metastasis: Mechanisms and Therapeutic Implications. Int J Mol Sci 2022; 23. doi: 10.3390/ijms23126806.
- Smeda M, Przyborowski K, Stojak M, Chlopicki S. The endothelial barrier and cancer metastasis: Does the protective facet of platelet function matter?. Biochem Pharmacol 2020; 176:113886. doi: 10.1016/j.bcp.2020.113886 [Crossref] [ Google Scholar]
- Rahman M, Mohammed S. Breast cancer metastasis and the lymphatic system. Oncol Lett 2015; 10:1233-9. doi: 10.3892/ol.2015.3486 [Crossref] [ Google Scholar]
-
Marconi GD, Fonticoli L, Rajan TS, Pierdomenico SD, Trubiani O, Pizzicannella J, et al. Epithelial-Mesenchymal Transition (EMT): The Type-2 EMT in Wound Healing, Tissue Regeneration and Organ Fibrosis. Cells 2021; 10. doi: 10.3390/cells10071587.
- Lamouille S, Xu J, Derynck R. Molecular mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell Biol 2014; 15:178-96. doi: 10.1038/nrm3758 [Crossref] [ Google Scholar]
-
Usman S, Waseem NH, Nguyen TKN, Mohsin S, Jamal A, Teh MT, et al. Vimentin Is at the Heart of Epithelial Mesenchymal Transition (EMT) Mediated Metastasis. Cancers (Basel) 2021; 13. doi: 10.3390/cancers13194985.
- Ilnitskaya AS, Litovka NI, Rubtsova SN, Zhitnyak IY, Gloushankova NA. Actin Cytoskeleton Remodeling Accompanied by Redistribution of Adhesion Proteins Drives Migration of Cells in Different EMT States. Cells 2024; 13. doi: 10.3390/cells13090780.
- Felding-Habermann B, O'Toole TE, Smith JW, Fransvea E, Ruggeri ZM, Ginsberg MH. Integrin activation controls metastasis in human breast cancer. Proc Natl Acad Sci U S A 2001; 98:1853-8. doi: 10.1073/pnas.98.4.1853 [Crossref] [ Google Scholar]
- Dai Y, Zhang X, Ou Y, Zou L, Zhang D, Yang Q. Anoikis resistance--protagonists of breast cancer cells survive and metastasize after ECM detachment. Cell Commun Signal 2023; 21:190. doi: 10.1186/s12964-023-01183-4 [Crossref] [ Google Scholar]
- Frisch SM, Schaller M, Cieply B. Mechanisms that link the oncogenic epithelial-mesenchymal transition to suppression of anoikis. J Cell Sci 2013; 126:21-9. doi: 10.1242/jcs.120907 [Crossref] [ Google Scholar]
- Garg M. Epithelial, mesenchymal and hybrid epithelial/mesenchymal phenotypes and their clinical relevance in cancer metastasis. Expert Rev Mol Med 2017; 19:e3. doi: 10.1017/erm.2017.6 [Crossref] [ Google Scholar]
- Zhu J, Wang R, Yang C, Shao X, Zhang Y, Hou J. Blocking tumor-platelet crosstalk to prevent tumor metastasis via reprograming glycolysis using biomimetic membrane-hybridized liposomes. J Control Release 2024; 366:328-41. doi: 10.1016/j.jconrel.2023.12.052 [Crossref] [ Google Scholar]
- Kopp HG, Placke T, Salih HR. Platelet-derived transforming growth factor-beta down-regulates NKG2D thereby inhibiting natural killer cell antitumor reactivity. Cancer Res 2009; 69:7775-83. doi: 10.1158/0008-5472.Can-09-2123 [Crossref] [ Google Scholar]
- Taftaf R, Liu X, Singh S, Jia Y, Dashzeveg NK, Hoffmann AD. ICAM1 initiates CTC cluster formation and trans-endothelial migration in lung metastasis of breast cancer. Nat Commun 2021; 12:4867. doi: 10.1038/s41467-021-25189-z [Crossref] [ Google Scholar]
- Gálvez BG, Matías-Román S, Albar JP, Sánchez-Madrid F, Arroyo AG. Membrane type 1-matrix metalloproteinase is activated during migration of human endothelial cells and modulates endothelial motility and matrix remodeling. J Biol Chem 2001; 276:37491-500. doi: 10.1074/jbc.M104094200 [Crossref] [ Google Scholar]
- Chao Y, Wu Q, Acquafondata M, Dhir R, Wells A. Partial mesenchymal to epithelial reverting transition in breast and prostate cancer metastases. Cancer Microenviron 2012; 5:19-28. doi: 10.1007/s12307-011-0085-4 [Crossref] [ Google Scholar]
-
Butturini E, Carcereri de Prati A, Boriero D, Mariotto S. Tumor Dormancy and Interplay with Hypoxic Tumor Microenvironment. Int J Mol Sci 2019; 20. doi: 10.3390/ijms20174305.
- Li M, Ding W, Sun T, Tariq MA, Xu T, Li P. Biogenesis of circular RNAs and their roles in cardiovascular development and pathology. Febs j 2018; 285:220-32. doi: 10.1111/febs.14191 [Crossref] [ Google Scholar]
- Yang L, Fu J, Zhou Y. Circular RNAs and Their Emerging Roles in Immune Regulation. Front Immunol 2018; 9:2977. doi: 10.3389/fimmu.2018.02977 [Crossref] [ Google Scholar]
- Ma Y, Zheng L, Gao Y, Zhang W, Zhang Q, Xu Y. A Comprehensive Overview of circRNAs: Emerging Biomarkers and Potential Therapeutics in Gynecological Cancers. Front Cell Dev Biol 2021; 9:709512. doi: 10.3389/fcell.2021.709512 [Crossref] [ Google Scholar]
- Schmidt CA, Giusto JD, Bao A, Hopper AK, Matera AG. Molecular determinants of metazoan tricRNA biogenesis. Nucleic Acids Res 2019; 47:6452-65. doi: 10.1093/nar/gkz311 [Crossref] [ Google Scholar]
- Zhang Y, Zhang XO, Chen T, Xiang JF, Yin QF, Xing YH. Circular intronic long noncoding RNAs. Mol Cell 2013; 51:792-806. doi: 10.1016/j.molcel.2013.08.017 [Crossref] [ Google Scholar]
-
Papatsirou M, Artemaki PI, Karousi P, Scorilas A, Kontos CK. Circular RNAs: Emerging Regulators of the Major Signaling Pathways Involved in Cancer Progression. Cancers (Basel) 2021; 13. doi: 10.3390/cancers13112744.
- Garlapati P, Ling J, Chiao PJ, Fu J. Circular RNAs regulate cancer-related signaling pathways and serve as potential diagnostic biomarkers for human cancers. Cancer Cell Int 2021; 21:317. doi: 10.1186/s12935-021-02017-4 [Crossref] [ Google Scholar]
- Hofmann S, Kedersha N, Anderson P, Ivanov P. Molecular mechanisms of stress granule assembly and disassembly. Biochim Biophys Acta Mol Cell Res 2021; 1868:118876. doi: 10.1016/j.bbamcr.2020.118876 [Crossref] [ Google Scholar]
- Wang F, Li J, Fan S, Jin Z, Huang C. Targeting stress granules: A novel therapeutic strategy for human diseases. Pharmacol Res 2020; 161:105143. doi: 10.1016/j.phrs.2020.105143 [Crossref] [ Google Scholar]
- Pfafenrot C, Preußer C. Establishing essential quality criteria for the validation of circular RNAs as biomarkers. Biomol Detect Quantif 2019; 17:100085. doi: 10.1016/j.bdq.2019.100085 [Crossref] [ Google Scholar]
- Dodbele S, Mutlu N, Wilusz JE. Best practices to ensure robust investigation of circular RNAs: pitfalls and tips. EMBO Rep 2021; 22:e52072. doi: 10.15252/embr.202052072 [Crossref] [ Google Scholar]
- Lin H, Conn VM, Conn SJ. Past, present, and future strategies for detecting and quantifying circular RNA variants. Febs j 2025; 292:4073-85. doi: 10.1111/febs.70012 [Crossref] [ Google Scholar]
- Vromman M, Yigit N, Verniers K, Lefever S, Vandesompele J, Volders PJ. Validation of Circular RNAs Using RT-qPCR After Effective Removal of Linear RNAs by Ribonuclease R. Curr Protoc 2021; 1:e181. doi: 10.1002/cpz1.181 [Crossref] [ Google Scholar]
- Schneider T, Schreiner S, Preußer C, Bindereif A, Rossbach O. Northern Blot Analysis of Circular RNAs. Methods Mol Biol 2018; 1724:119-33. doi: 10.1007/978-1-4939-7562-4_10 [Crossref] [ Google Scholar]
- Hansen TB, Venø MT, Damgaard CK, Kjems J. Comparison of circular RNA prediction tools. Nucleic Acids Res 2016; 44:e58. doi: 10.1093/nar/gkv1458 [Crossref] [ Google Scholar]
- Nielsen AF, Bindereif A, Bozzoni I, Hanan M, Hansen TB, Irimia M. Best practice standards for circular RNA research. Nat Methods 2022; 19:1208-20. doi: 10.1038/s41592-022-01487-2 [Crossref] [ Google Scholar]
- Niu M, Wang C, Chen Y, Zou Q, Qi R, Xu L. CircRNA identification and feature interpretability analysis. BMC Biol 2024; 22:44. doi: 10.1186/s12915-023-01804-x [Crossref] [ Google Scholar]
- Azizzad Ranji E, Kahrizi D, Khanahmadi M, Rashidi Monfared S. CircRNAs: Biogenesis, identification and expression analysis. Cellular, Molecular and Biomedical Reports 2025; 5:63-79. doi: 10.55705/cmbr.2025.460043.1248 [Crossref] [ Google Scholar]
- Naser Al Deen N, Atallah Lanman N, Chittiboyina S, Lelièvre S, Nasr R, Nassar F. A risk progression breast epithelial 3D culture model reveals Cx43/hsa_circ_0077755/miR-182 as a biomarker axis for heightened risk of breast cancer initiation. Sci Rep 2021; 11:2626. doi: 10.1038/s41598-021-82057-y [Crossref] [ Google Scholar]
- Zhang W, Xie X, Huang Z, Zhong X, Liu Y, Cheong KL. The integration of single-cell sequencing, TCGA, and GEO data analysis revealed that PRRT3-AS1 is a biomarker and therapeutic target of SKCM. Front Immunol 2022; 13:919145. doi: 10.3389/fimmu.2022.919145 [Crossref] [ Google Scholar]
- Hawsawi YM, Khoja B, Aljaylani AO, Jaha R, AlDerbi RM, Alnuman H. Recent progress and applications of single-cell sequencing technology in breast cancer. Front Genet 2024; 15:1417415. doi: 10.3389/fgene.2024.1417415 [Crossref] [ Google Scholar]
- Savagner P, Gilles C, Newgreen DF, Sato H, Thompson EW. Matrix metalloproteases and epithelial-to-mesenchymal transition: implications for carcinoma metastasis. Rise Fall Epithelial Phenotype 2005; 297-315.
-
Datta A, Deng S, Gopal V, Yap KC, Halim CE, Lye ML, et al. Cytoskeletal Dynamics in Epithelial-Mesenchymal Transition: Insights into Therapeutic Targets for Cancer Metastasis. Cancers (Basel) 2021; 13. doi: 10.3390/cancers13081882.
- Leggett SE, Hruska AM, Guo M, Wong IY. The epithelial-mesenchymal transition and the cytoskeleton in bioengineered systems. Cell Commun Signal 2021; 19:32. doi: 10.1186/s12964-021-00713-2 [Crossref] [ Google Scholar]
- Pradeau-Phélut L, Etienne-Manneville S. Cytoskeletal crosstalk: A focus on intermediate filaments. Curr Opin Cell Biol 2024; 87:102325. doi: 10.1016/j.ceb.2024.102325 [Crossref] [ Google Scholar]
- Asadi A, Molavand M, Vaghari-Tabari M, Yousefi B. Wnt ligands in breast cancer metastasis: unraveling their dual roles and therapeutic potential. Mol Biol Rep 2026; 53:280. doi: 10.1007/s11033-026-11431-8 [Crossref] [ Google Scholar]
- Vora HH, Patel NA, Rajvik KN, Mehta SV, Brahmbhatt BV, Shah MJ. Cytokeratin and vimentin expression in breast cancer. Int J Biol Markers 2009; 24:38-46. doi: 10.1177/172460080902400106 [Crossref] [ Google Scholar]
- Vallenius T. Actin stress fibre subtypes in mesenchymal-migrating cells. Open Biol 2013; 3:130001. doi: 10.1098/rsob.130001 [Crossref] [ Google Scholar]
- Nurmagambetova A, Mustyatsa V, Saidova A, Vorobjev I. Morphological and cytoskeleton changes in cells after EMT. Sci Rep 2023; 13:22164. doi: 10.1038/s41598-023-48279-y [Crossref] [ Google Scholar]
-
Fu A, Yao B, Dong T, Chen Y, Yao J, Liu Y, et al. Tumor-resident intracellular microbiota promotes metastatic colonization in breast cancer. Cell 2022; 185: 1356-1372.e26. doi: 10.1016/j.cell.2022.02.027.
- Klubíčková N. The role of microtubule dynamics in cancer metastasis [thesis]. Univerzita Karlova, Přírodovědecká fakulta; 2020.
- Enderle-Ammour K, Bader M, Ahrens TD, Franke K, Timme S, Csanadi A. Form follows function: Morphological and immunohistological insights into epithelial-mesenchymal transition characteristics of tumor buds. Tumour Biol 2017; 39:1010428317705501. doi: 10.1177/1010428317705501 [Crossref] [ Google Scholar]
- Baba AB, Rah B, Bhat GR, Mushtaq I, Parveen S, Hassan R. Transforming Growth Factor-Beta (TGF-β) Signaling in Cancer-A Betrayal Within. Front Pharmacol 2022; 13:791272. doi: 10.3389/fphar.2022.791272 [Crossref] [ Google Scholar]
-
Heldin CH, Moustakas A. Signaling Receptors for TGF-β Family Members. Cold Spring Harb Perspect Biol 2016; 8. doi: 10.1101/cshperspect.a022053.
- Tang LY, Heller M, Meng Z, Yu LR, Tang Y, Zhou M. Transforming Growth Factor-β (TGF-β) Directly Activates the JAK1-STAT3 Axis to Induce Hepatic Fibrosis in Coordination with the SMAD Pathway. J Biol Chem 2017; 292:4302-12. doi: 10.1074/jbc.M116.773085 [Crossref] [ Google Scholar]
-
Hata A, Chen YG. TGF-β Signaling from Receptors to Smads. Cold Spring Harb Perspect Biol 2016; 8. doi: 10.1101/cshperspect.a022061.
- Howe PH. CHAPTER 49 - Transforming growth factor β. In: Thomson AW, MT Lotze, editors. The Cytokine Handbook (Fourth Edition). London: Academic Press. 2003. p. 1119-52d.
- Tarasewicz E, Jeruss JS. Phospho-specific Smad3 signaling: impact on breast oncogenesis. Cell Cycle 2012; 11:2443-51. doi: 10.4161/cc.20546 [Crossref] [ Google Scholar]
- Moustakas A, Souchelnytskyi S, Heldin CH. Smad regulation in TGF-beta signal transduction. J Cell Sci 2001; 114:4359-69. doi: 10.1242/jcs.114.24.4359 [Crossref] [ Google Scholar]
- Nakao A, Imamura T, Souchelnytskyi S, Kawabata M, Ishisaki A, Oeda E. TGF-beta receptor-mediated signalling through Smad2, Smad3 and Smad4. Embo j 1997; 16:5353-62. doi: 10.1093/emboj/16.17.5353 [Crossref] [ Google Scholar]
- Song B, Estrada KD, Lyons KM. Smad signaling in skeletal development and regeneration. Cytokine Growth Factor Rev 2009; 20:379-88. doi: 10.1016/j.cytogfr.2009.10.010 [Crossref] [ Google Scholar]
- Fuxe J, Vincent T, Garcia de Herreros A. Transcriptional crosstalk between TGF-β and stem cell pathways in tumor cell invasion: role of EMT promoting Smad complexes. Cell Cycle 2010; 9:2363-74. doi: 10.4161/cc.9.12.12050 [Crossref] [ Google Scholar]
- Zeng K, He B, Yang BB, Xu T, Chen X, Xu M. The pro-metastasis effect of circANKS1B in breast cancer. Mol Cancer 2018; 17:160. doi: 10.1186/s12943-018-0914-x [Crossref] [ Google Scholar]
- Ren YQ, Li QH, Liu LB. USF1 prompt melanoma through upregulating TGF-β signaling pathway. Eur Rev Med Pharmacol Sci 2016; 20:3592-8. [ Google Scholar]
- Leng X, Huang G, Ding J, Ma F. Circ_0000043 promotes the proliferation, migration, invasiveness, and epithelial-mesenchymal transition in breast cancer cells via the miR-136-Smad3 axis. Biochem Cell Biol 2021; 99:277-85. doi: 10.1139/bcb-2020-0219 [Crossref] [ Google Scholar]
- Li H, Xue S, Zhang X, Li F, Bei S, Feng L. CircRNA PVT1 modulated cell migration and invasion through Epithelial-Mesenchymal Transition (EMT) mediation in gastric cancer through miR-423-5p/Smad3 pathway. Regen Ther 2022; 21:25-33. doi: 10.1016/j.reth.2022.02.003 [Crossref] [ Google Scholar]
- Liu J, Dai X, Guo X, Cheng A, Mac SM, Wang Z. Circ-OXCT1 Suppresses Gastric Cancer EMT and Metastasis by Attenuating TGF-β Pathway Through the Circ-OXCT1/miR-136/SMAD4 Axis. Onco Targets Ther 2020; 13:3987-98. doi: 10.2147/ott.S239789 [Crossref] [ Google Scholar]
- Wang L, Tong X, Zhou Z, Wang S, Lei Z, Zhang T. Circular RNA hsa_circ_0008305 (circPTK2) inhibits TGF-β-induced epithelial-mesenchymal transition and metastasis by controlling TIF1γ in non-small cell lung cancer. Mol Cancer 2018; 17:140. doi: 10.1186/s12943-018-0889-7 [Crossref] [ Google Scholar]
- Zhu J, Luo Y, Zhao Y, Kong Y, Zheng H, Li Y. circEHBP1 promotes lymphangiogenesis and lymphatic metastasis of bladder cancer via miR-130a-3p/TGFβR1/VEGF-D signaling. Mol Ther 2021; 29:1838-52. doi: 10.1016/j.ymthe.2021.01.031 [Crossref] [ Google Scholar]
- Zheng L, Liang H, Zhang Q, Shen Z, Sun Y, Zhao X. circPTEN1, a circular RNA generated from PTEN, suppresses cancer progression through inhibition of TGF-β/Smad signaling. Mol Cancer 2022; 21:41. doi: 10.1186/s12943-022-01495-y [Crossref] [ Google Scholar]
- Liu J, Xiao Q, Xiao J, Niu C, Li Y, Zhang X. Wnt/β-catenin signalling: function, biological mechanisms, and therapeutic opportunities. Signal Transduct Target Ther 2022; 7:3. doi: 10.1038/s41392-021-00762-6 [Crossref] [ Google Scholar]
- Molavand M, Majidinia M, Yousefi B. Critical Contribution of Various Signaling Pathways in the Development of Drug Resistance in Colorectal Cancer: An Update. IUBMB Life 2025; 77:e70074. doi: 10.1002/iub.70074 [Crossref] [ Google Scholar]
- Akiyama T. Wnt/beta-catenin signaling. Cytokine Growth Factor Rev 2000; 11:273-82. doi: 10.1016/s1359-6101(00)00011-3 [Crossref] [ Google Scholar]
- MacDonald BT, Tamai K, He X. Wnt/beta-catenin signaling: components, mechanisms, and diseases. Dev Cell 2009; 17:9-26. doi: 10.1016/j.devcel.2009.06.016 [Crossref] [ Google Scholar]
-
Marineau A, Khan KA, Servant MJ. Roles of GSK-3 and β-Catenin in Antiviral Innate Immune Sensing of Nucleic Acids. Cells 2020; 9. doi: 10.3390/cells9040897.
-
Shishido A, Miyo M, Oishi K, Nishiyama N, Wu M, Yamamoto H, et al. The Relationship between LRP6 and Wnt/β-Catenin Pathway in Colorectal and Esophageal Cancer. Life (Basel) 2023; 13. doi: 10.3390/life13030615.
- Ren Q, Chen J, Liu Y. LRP5 and LRP6 in Wnt Signaling: Similarity and Divergence. Front Cell Dev Biol 2021; 9:670960. doi: 10.3389/fcell.2021.670960 [Crossref] [ Google Scholar]
- Zhao Z, Han X, Nie C, Lin S, Wang J, Fang H. Circ_0008784 activates Wnt/β-catenin pathway to affect the proliferation and apoptosis of triple-negative breast cancer cells. Pathol Res Pract 2023; 241:154185. doi: 10.1016/j.prp.2022.154185 [Crossref] [ Google Scholar]
- Wang ST, Liu LB, Li XM, Wang YF, Xie PJ, Li Q. Circ-ITCH regulates triple-negative breast cancer progression through the Wnt/β-catenin pathway. Neoplasma 2019; 66:232-9. doi: 10.4149/neo_2018_180710N460 [Crossref] [ Google Scholar]
- Jiang J, Cheng X. Circular RNA circABCC4 acts as a ceRNA of miR-154-5p to improve cell viability, migration and invasion of breast cancer cells in vitro. Cell Cycle 2020; 19:2653-61. doi: 10.1080/15384101.2020.1815147 [Crossref] [ Google Scholar]
- Zhang J, Xu HD, Xing XJ, Liang ZT, Xia ZH, Zhao Y. CircRNA_069718 promotes cell proliferation and invasion in triple-negative breast cancer by activating Wnt/β-catenin pathway. Eur Rev Med Pharmacol Sci 2019; 23:5315-22. doi: 10.26355/eurrev_201906_18198 [Crossref] [ Google Scholar]
- Yang SJ, Wang DD, Zhong SL, Chen WQ, Wang FL, Zhang J. Tumor-derived exosomal circPSMA1 facilitates the tumorigenesis, metastasis, and migration in triple-negative breast cancer (TNBC) through miR-637/Akt1/β-catenin (cyclin D1) axis. Cell Death Dis 2021; 12:420. doi: 10.1038/s41419-021-03680-1 [Crossref] [ Google Scholar]
- Li Y, Wang Z, Su P, Liang Y, Li Z, Zhang H. circ-EIF6 encodes EIF6-224aa to promote TNBC progression via stabilizing MYH9 and activating the Wnt/beta-catenin pathway. Mol Ther 2022; 30:415-30. doi: 10.1016/j.ymthe.2021.08.026 [Crossref] [ Google Scholar]
- Liu X, Song J, Kang Y, Wang Y, Chen A. CircPDSS1 promotes the proliferation, invasion, migration, and EMT of breast cancer cell via regulating miR-320c/CKAP5 axis. Cancer Cell Int 2022; 22:238. doi: 10.1186/s12935-022-02657-0 [Crossref] [ Google Scholar]
- Zhang X, Yao J, Shi H, Gao B, Zhou H, Zhang Y. Hsa_circ_0026628 promotes the development of colorectal cancer by targeting SP1 to activate the Wnt/β-catenin pathway. Cell Death Dis 2021; 12:802. doi: 10.1038/s41419-021-03794-6 [Crossref] [ Google Scholar]
- Shen T, Cheng X, Liu X, Xia C, Zhang H, Pan D. Circ_0026344 restrains metastasis of human colorectal cancer cells via miR-183. Artif Cells Nanomed Biotechnol 2019; 47:4038-45. doi: 10.1080/21691401.2019.1669620 [Crossref] [ Google Scholar]
- Gu H, Xu Z, Zhang J, Wei Y, Cheng L, Wang J. circ_0038718 promotes colon cancer cell malignant progression via the miR-195-5p/Axin2 signaling axis and also effect Wnt/β-catenin signal pathway. BMC Genomics 2021; 22:768. doi: 10.1186/s12864-021-07880-z [Crossref] [ Google Scholar]
- Li XY, Liu YR, Zhou JH, Li W, Guo HH, Ma HP. Enhanced expression of circular RNA hsa_circ_000984 promotes cells proliferation and metastasis in non-small cell lung cancer by modulating Wnt/β-catenin pathway. Eur Rev Med Pharmacol Sci 2019; 23:3366-74. doi: 10.26355/eurrev_201904_17700 [Crossref] [ Google Scholar]
- Gao S, Yu Y, Liu L, Meng J, Li G. Circular RNA hsa_circ_0007059 restrains proliferation and epithelial-mesenchymal transition in lung cancer cells via inhibiting microRNA-378. Life Sci 2019; 233:116692. doi: 10.1016/j.lfs.2019.116692 [Crossref] [ Google Scholar]
- Fresno Vara JA, Casado E, de Castro J, Cejas P, Belda-Iniesta C, González-Barón M. PI3K/Akt signalling pathway and cancer. Cancer Treat Rev 2004; 30:193-204. doi: 10.1016/j.ctrv.2003.07.007 [Crossref] [ Google Scholar]
- Manning BD, Toker A. AKT/PKB Signaling: Navigating the Network. Cell 2017; 169:381-405. doi: 10.1016/j.cell.2017.04.001 [Crossref] [ Google Scholar]
- Chen W, Wu S, Zhang G, Wang W, Shi Y. Effect of AKT inhibition on epithelial-mesenchymal transition and ZEB1-potentiated radiotherapy in nasopharyngeal carcinoma. Oncol Lett 2013; 6:1234-40. doi: 10.3892/ol.2013.1552 [Crossref] [ Google Scholar]
- Orellana AM, Vasconcelos AR, Leite JA, de Sá Lima L, Andreotti DZ, Munhoz CD. Age-related neuroinflammation and changes in AKT-GSK-3β and WNT/ β-CATENIN signaling in rat hippocampus. Aging (Albany NY) 2015; 7:1094-111. doi: 10.18632/aging.100853 [Crossref] [ Google Scholar]
- Xu JH, Wang Y, Xu D. Hsa_circ_001569 is an unfavorable prognostic factor and promotes cell proliferation and metastasis by modulating PI3K-AKT pathway in breast cancer. Cancer Biomark 2019; 25:193-201. doi: 10.3233/cbm-182293 [Crossref] [ Google Scholar]
- Wang X, Jian W, Luo Q, Fang L. CircSEMA4B inhibits the progression of breast cancer by encoding a novel protein SEMA4B-211aa and regulating AKT phosphorylation. Cell Death Dis 2022; 13:794. doi: 10.1038/s41419-022-05246-1 [Crossref] [ Google Scholar]
- He J, Wang J, Li S, Li T, Chen K, Zhang S. Hypoxia-inhibited miR-338-3p suppresses breast cancer progression by directly targeting ZEB2. Cancer Sci 2020; 111:3550-63. doi: 10.1111/cas.14589 [Crossref] [ Google Scholar]
- Tang H, Huang X, Wang J, Yang L, Kong Y, Gao G. circKIF4A acts as a prognostic factor and mediator to regulate the progression of triple-negative breast cancer. Mol Cancer 2019; 18:23. doi: 10.1186/s12943-019-0946-x [Crossref] [ Google Scholar]
- Zhou J, Song S, He S, Zhu X, Zhang Y, Yi B. MicroRNA-375 targets PDK1 in pancreatic carcinoma and suppresses cell growth through the Akt signaling pathway. Int J Mol Med 2014; 33:950-6. doi: 10.3892/ijmm.2014.1638 [Crossref] [ Google Scholar]
- Chen Y, Wang D, Shu T, Sun K, Zhao J, Wang M. Circular RNA_0000326 promotes bladder cancer progression via microRNA-338-3p/ETS Proto-Oncogene 1/phosphoinositide-3 kinase/Akt pathway. Bioengineered 2021; 12:11410-22. doi: 10.1080/21655979.2021.2008738 [Crossref] [ Google Scholar]
- Xu G, Zhang P, Liang H, Xu Y, Shen J, Wang W. Circular RNA hsa_circ_0003288 induces EMT and invasion by regulating hsa_circ_0003288/miR-145/PD-L1 axis in hepatocellular carcinoma. Cancer Cell Int 2021; 21:212. doi: 10.1186/s12935-021-01902-2 [Crossref] [ Google Scholar]
- Zhu Q, Zhang X, Zai HY, Jiang W, Zhang KJ, He YQ. circSLC8A1 sponges miR-671 to regulate breast cancer tumorigenesis via PTEN/PI3k/Akt pathway. Genomics 2021; 113:398-410. doi: 10.1016/j.ygeno.2020.12.006 [Crossref] [ Google Scholar]
- Zhang XY, Mao L. Circular RNA Circ_0000442 acts as a sponge of MiR-148b-3p to suppress breast cancer via PTEN/PI3K/Akt signaling pathway. Gene 2021; 766:145113. doi: 10.1016/j.gene.2020.145113 [Crossref] [ Google Scholar]
- Zhou X, Jian W, Luo Q, Zheng W, Deng X, Wang X. Circular RNA_0006014 promotes breast cancer progression through sponging miR-885-3p to regulate NTRK2 and PIK3/AKT pathway. Aging (Albany NY) 2022; 14:3105-28. doi: 10.18632/aging.203996 [Crossref] [ Google Scholar]
- Ji C, Zhu L, Fang L. Hsa_circ_0000851 promotes PDK1/p-AKT-mediated cell proliferation and migration by regulating miR-1183 in triple-negative breast cancer. Cell Signal 2023; 101:110494. doi: 10.1016/j.cellsig.2022.110494 [Crossref] [ Google Scholar]
- Wang L, Zhou Y, Jiang L, Lu L, Dai T, Li A. CircWAC induces chemotherapeutic resistance in triple-negative breast cancer by targeting miR-142, upregulating WWP1 and activating the PI3K/AKT pathway. Mol Cancer 2021; 20:43. doi: 10.1186/s12943-021-01332-8 [Crossref] [ Google Scholar]
- Cheng Z, Liu G, Huang C, Zhao X. Upregulation of circRNA_100395 sponges miR-142-3p to inhibit gastric cancer progression by targeting the PI3K/AKT axis. Oncol Lett 2021; 21:419. doi: 10.3892/ol.2021.12680 [Crossref] [ Google Scholar]
- Jiang X, Sorkin A. Coordinated traffic of Grb2 and Ras during epidermal growth factor receptor endocytosis visualized in living cells. Mol Biol Cell 2002; 13:1522-35. doi: 10.1091/mbc.01-11-0552 [Crossref] [ Google Scholar]
- Cseh B, Doma E, Baccarini M. "RAF" neighborhood: protein-protein interaction in the Raf/Mek/Erk pathway. FEBS Lett 2014; 588:2398-406. doi: 10.1016/j.febslet.2014.06.025 [Crossref] [ Google Scholar]
- Chapnick DA, Warner L, Bernet J, Rao T, Liu X. Partners in crime: the TGFβ and MAPK pathways in cancer progression. Cell Biosci 2011; 1:42. doi: 10.1186/2045-3701-1-42 [Crossref] [ Google Scholar]
- Hua K, Gong H, Xu Q, Li T, Ma B, Li Y. P38 mitogen-activated protein kinase promotes Wnt/β-catenin signaling by impeding Dickkofp-1 expression during Haemophilus parasuis infection. Cytokine 2020; 136:155287. doi: 10.1016/j.cyto.2020.155287 [Crossref] [ Google Scholar]
- Kar R, Jha NK, Jha SK, Sharma A, Dholpuria S, Asthana N. A "NOTCH" Deeper into the Epithelial-To-Mesenchymal Transition (EMT) Program in Breast Cancer. Genes (Basel) 2019; 10:961. doi: 10.3390/genes10120961 [Crossref] [ Google Scholar]
- Chen H, Wang X, Cheng H, Deng Y, Chen J, Wang B. CircRNA circRREB1 promotes tumorigenesis and progression of breast cancer by activating Erk1/2 signaling through interacting with GNB4. Heliyon 2024; 10:e28785. doi: 10.1016/j.heliyon.2024.e28785 [Crossref] [ Google Scholar]
- Zhang Y, Li Y, Wang Q, Su B, Xu H, Sun Y. Role of RASA1 in cancer: A review and update (Review). Oncol Rep 2020; 44:2386-96. doi: 10.3892/or.2020.7807 [Crossref] [ Google Scholar]
- Xiao W, Zheng S, Zou Y, Yang A, Xie X, Tang H. CircAHNAK1 inhibits proliferation and metastasis of triple-negative breast cancer by modulating miR-421 and RASA1. Aging (Albany NY) 2019; 11:12043-56. doi: 10.18632/aging.102539 [Crossref] [ Google Scholar]
- Chen Q, Yang Z, Ding H, Li H, Wang W, Pan Z. CircWHSC1 Promotes Breast Cancer Progression by Regulating the FASN/AMPK/mTOR Axis Through Sponging miR-195-5p. Front Oncol 2021; 11:649242. doi: 10.3389/fonc.2021.649242 [Crossref] [ Google Scholar]
- Ju C, Zhou M, Du D, Wang C, Yao J, Li H. EIF4A3-mediated circ_0042881 activates the RAS pathway via miR-217/SOS1 axis to facilitate breast cancer progression. Cell Death Dis 2023; 14:559. doi: 10.1038/s41419-023-06085-4 [Crossref] [ Google Scholar]
- Wang Y, Xu R, Zhang D, Lu T, Yu W, Wo Y. Circ-ZKSCAN1 regulates FAM83A expression and inactivates MAPK signaling by targeting miR-330-5p to promote non-small cell lung cancer progression. Transl Lung Cancer Res 2019; 8:862-75. doi: 10.21037/tlcr.2019.11.04 [Crossref] [ Google Scholar]
- Zhang X, Wang S, Wang H, Cao J, Huang X, Chen Z. Circular RNA circNRIP1 acts as a microRNA-149-5p sponge to promote gastric cancer progression via the AKT1/mTOR pathway. Mol Cancer 2019; 18:20. doi: 10.1186/s12943-018-0935-5 [Crossref] [ Google Scholar]
- Li X, Ma N, Zhang Y, Wei H, Zhang H, Pang X. Circular RNA circNRIP1 promotes migration and invasion in cervical cancer by sponging miR-629-3p and regulating the PTP4A1/ERK1/2 pathway. Cell Death Dis 2020; 11:399. doi: 10.1038/s41419-020-2607-9 [Crossref] [ Google Scholar]
- Khoshbakht S, Zomorodi Anbaji F, Darzi M, Esmaeili R. The endogenous association among MMP2/miR-1248/Circ_0087558/miR-643/ MAP2K6 axis can contribute to brain metastasis in basal-like subtype of breast cancer. Heliyon 2024; 10:e33195. doi: 10.1016/j.heliyon.2024.e33195 [Crossref] [ Google Scholar]
- Gu Y, Mohammad IS, Liu Z. Overview of the STAT-3 signaling pathway in cancer and the development of specific inhibitors. Oncol Lett 2020; 19:2585-94. doi: 10.3892/ol.2020.11394 [Crossref] [ Google Scholar]
- Liu YP, Heng JY, Zhao XY, Li EY. The inhibition of circular RNA circNOLC1 by propofol/STAT3 attenuates breast cancer stem cells function via miR-365a-3p/STAT3 signaling. J Transl Med 2021; 19:467. doi: 10.1186/s12967-021-03133-5 [Crossref] [ Google Scholar]
- Zheng X, Huang M, Xing L, Yang R, Wang X, Jiang R. The circRNA circSEPT9 mediated by E2F1 and EIF4A3 facilitates the carcinogenesis and development of triple-negative breast cancer. Mol Cancer 2020; 19:73. doi: 10.1186/s12943-020-01183-9 [Crossref] [ Google Scholar]
- Min J, Pan X, Lv G. The circRNA circ_0000291 acts as a sponge of microRNA 326 to regulate E26 transformation-specific sequence-1 expression and promote breast cancer progression. Pathol Int 2020; 70:953-64. doi: 10.1111/pin.13011 [Crossref] [ Google Scholar]
- Chen G, Shi Y, Zhang Y, Sun J. CircRNA_100782 regulates pancreatic carcinoma proliferation through the IL6-STAT3 pathway. Onco Targets Ther 2017; 10:5783-94. doi: 10.2147/ott.S150678 [Crossref] [ Google Scholar]
- Liu L, Liu FB, Huang M, Xie K, Xie QS, Liu CH. Circular RNA ciRS-7 promotes the proliferation and metastasis of pancreatic cancer by regulating miR-7-mediated EGFR/STAT3 signaling pathway. Hepatobiliary Pancreat Dis Int 2019; 18:580-6. doi: 10.1016/j.hbpd.2019.03.003 [Crossref] [ Google Scholar]
- Zhang Z, Sun C, Zheng Y, Gong Y. circFCHO2 promotes gastric cancer progression by activating the JAK1/STAT3 pathway via sponging miR-194-5p. Cell Cycle 2022; 21:2145-64. doi: 10.1080/15384101.2022.2087280 [Crossref] [ Google Scholar]
- Liu J, Xie J, Xu E, Xu B, Zhou J, Zhou J. CircRNA hsa_circ_0000043 acts as a miR-4492 sponge to promote lung cancer progression via BDNF and STAT3 expression regulation in anti-benzo[a]pyrene-trans-7,8-dihydrodiol-9,10-epoxide-transformed 16HBE cells. Toxicol Sci 2023; 195:87-102. doi: 10.1093/toxsci/kfad060 [Crossref] [ Google Scholar]
- Chen Z, Cheng H, Zhang J, Jiang D, Chen G, Yan S. Hsa_circRNA_102051 regulates colorectal cancer proliferation and metastasis by mediating Notch pathway. Cancer Cell Int 2023; 23:230. doi: 10.1186/s12935-023-03026-1 [Crossref] [ Google Scholar]
- Shi YR, Wu Z, Xiong K, Liao QJ, Ye X, Yang P. Circular RNA circKIF4A Sponges miR-375/1231 to Promote Bladder Cancer Progression by Upregulating NOTCH2 Expression. Front Pharmacol 2020; 11:605. doi: 10.3389/fphar.2020.00605 [Crossref] [ Google Scholar]
- Wu HB, Huang SS, Lu CG, Tian SD, Chen M. CircAPLP2 regulates the proliferation and metastasis of colorectal cancer by targeting miR-101-3p to activate the Notch signalling pathway. Am J Transl Res 2020; 12:2554-69. [ Google Scholar]
- Huang Z, Ding Y, Zhang L, He S, Jia Z, Gu C. Upregulated circPDK1 Promotes RCC Cell Migration and Invasion by Regulating the miR-377-3P-NOTCH1 Axis in Renal Cell Carcinoma. Onco Targets Ther 2020; 13:11237-52. doi: 10.2147/ott.S280434 [Crossref] [ Google Scholar]
- Tang J, Huang M, Peng H, He J, Gong L, Yang M. METTL3-mediated Hsa_circ_0000390 downregulation enhances the proliferation, migration, and invasion of colorectal cancer cells by increasing Notch1 expression. Hum Cell 2023; 36:703-11. doi: 10.1007/s13577-022-00835-w [Crossref] [ Google Scholar]
- He Y, Huang H, Jin L, Zhang F, Zeng M, Wei L. CircZNF609 enhances hepatocellular carcinoma cell proliferation, metastasis, and stemness by activating the Hedgehog pathway through the regulation of miR-15a-5p/15b-5p and GLI2 expressions. Cell Death Dis 2020; 11:358. doi: 10.1038/s41419-020-2441-0 [Crossref] [ Google Scholar]
- Jiang Y, Xiao F, Wang L, Wang T, Chen L. Circular RNA has_circ_0000034 accelerates retinoblastoma advancement through the miR-361-3p/ADAM19 axis. Mol Cell Biochem 2021; 476:69-80. doi: 10.1007/s11010-020-03886-5 [Crossref] [ Google Scholar]
- Zhao B, Song X, Guan H. CircACAP2 promotes breast cancer proliferation and metastasis by targeting miR-29a/b-3p-COL5A1 axis. Life Sci 2020; 244:117179. doi: 10.1016/j.lfs.2019.117179 [Crossref] [ Google Scholar]
- Qu Y, Dou P, Hu M, Xu J, Xia W, Sun H. circRNA-CER mediates malignant progression of breast cancer through targeting the miR-136/MMP13 axis. Mol Med Rep 2019; 19:3314-20. doi: 10.3892/mmr.2019.9965 [Crossref] [ Google Scholar]
- Du WW, Yang W, Li X, Fang L, Wu N, Li F. The Circular RNA circSKA3 Binds Integrin β1 to Induce Invadopodium Formation Enhancing Breast Cancer Invasion. Mol Ther 2020; 28:1287-98. doi: 10.1016/j.ymthe.2020.03.002 [Crossref] [ Google Scholar]
- Zhang HD, Jiang LH, Hou JC, Zhong SL, Zhou SY, Zhu LP. Circular RNA hsa_circ_0052112 promotes cell migration and invasion by acting as sponge for miR-125a-5p in breast cancer. Biomed Pharmacother 2018; 107:1342-53. doi: 10.1016/j.biopha.2018.08.030 [Crossref] [ Google Scholar]
- Yang L, Song C, Chen Y, Jing G, Sun J. Circular RNA circ_0103552 forecasts dismal prognosis and promotes breast cancer cell proliferation and invasion by sponging miR-1236. J Cell Biochem 2019; 120:15553-60. doi: 10.1002/jcb.28822 [Crossref] [ Google Scholar]
- Hou JC, Xu Z, Zhong SL, Zhang HD, Jiang LH, Chen X. Circular RNA circASS1 is downregulated in breast cancer cells MDA-MB-231 and suppressed invasion and migration. Epigenomics 2019; 11:199-213. doi: 10.2217/epi-2017-0167 [Crossref] [ Google Scholar]
- Li P, Ren X, Zheng Y, Sun J, Ye G. Tumor promoting effect of circ_002172 associates with induced immune escape in breast cancer via the miR-296-5p/CXCL12 axis. Int Immunopharmacol 2022; 106:108530. doi: 10.1016/j.intimp.2022.108530 [Crossref] [ Google Scholar]
- Zheng Y, Ren S, Zhang Y, Liu S, Meng L, Liu F. Circular RNA circWWC3 augments breast cancer progression through promoting M2 macrophage polarization and tumor immune escape via regulating the expression and secretion of IL-4. Cancer Cell Int 2022; 22:264. doi: 10.1186/s12935-022-02686-9 [Crossref] [ Google Scholar]
- Li J, Dong X, Kong X, Wang Y, Li Y, Tong Y. Circular RNA hsa_circ_0067842 facilitates tumor metastasis and immune escape in breast cancer through HuR/CMTM6/PD-L1 axis. Biol Direct 2023; 18:48. doi: 10.1186/s13062-023-00397-3 [Crossref] [ Google Scholar]
- Huang L, Ma J, Cui M. Circular RNA hsa_circ_0001598 promotes programmed death-ligand-1-mediated immune escape and trastuzumab resistance via sponging miR-1184 in breast cancer cells. Immunol Res 2021; 69:558-67. doi: 10.1007/s12026-021-09237-w [Crossref] [ Google Scholar]
-
Dong LF, Chen FF, Fan YF, Zhang K, Chen HH. circ-0000512 inhibits PD-L1 ubiquitination through sponging miR-622/CMTM6 axis to promote triple-negative breast cancer and immune escape. J Immunother Cancer 2023; 11. doi: 10.1136/jitc-2022-005461.
- Zhang Z, Huo W, Li J. circATAD2 mitigates CD8( + ) T cells antitumor immune surveillance in breast cancer via IGF2BP3/m(6)A/PD-L1 manner. Cancer Immunol Immunother 2024; 73:130. doi: 10.1007/s00262-024-03705-6 [Crossref] [ Google Scholar]
- Liang L, Gao M, Li W, Tang J, He Q, Zeng F. CircGSK3β mediates PD-L1 transcription through miR-338-3p/PRMT5/H3K4me3 to promote breast cancer cell immune evasion and tumor progression. Cell Death Discov 2024; 10:426. doi: 10.1038/s41420-024-02197-8 [Crossref] [ Google Scholar]
- Zetter BR. Angiogenesis and tumor metastasis. Annu Rev Med 1998; 49:407-24. doi: 10.1146/annurev.med.49.1.407 [Crossref] [ Google Scholar]
- Liu T, Ye P, Ye Y, Lu S, Han B. Circular RNA hsa_circRNA_002178 silencing retards breast cancer progression via microRNA-328-3p-mediated inhibition of COL1A1. J Cell Mol Med 2020; 24:2189-201. doi: 10.1111/jcmm.14875 [Crossref] [ Google Scholar]
- Chen Y, Zhou Y, Chen J, Yang J, Yuan Y, Wu W. Exosomal lncRNA SNHG12 promotes angiogenesis and breast cancer progression. Breast Cancer 2024; 31:607-20. doi: 10.1007/s12282-024-01574-6 [Crossref] [ Google Scholar]
- Su M, Xiao Y, Ma J, Tang Y, Tian B, Zhang Y. Circular RNAs in Cancer: emerging functions in hallmarks, stemness, resistance and roles as potential biomarkers. Mol Cancer 2019; 18:90. doi: 10.1186/s12943-019-1002-6 [Crossref] [ Google Scholar]
- Carlos-Reyes Á, Romero-Garcia S, Contreras-Sanzón E, Ruiz V, Prado-Garcia H. Role of Circular RNAs in the Regulation of Immune Cells in Response to Cancer Therapies. Front Genet 2022; 13:823238. doi: 10.3389/fgene.2022.823238 [Crossref] [ Google Scholar]
-
De Palma FD, Salvatore F, Pol JG, Kroemer G, Maiuri MC. Circular RNAs as Potential Biomarkers in Breast Cancer. Biomedicines [serial on the Internet] 2022; 10(3). doi: 10.3390/biomedicines10030725.
- Darbeheshti F, Zokaei E, Mansoori Y, Emadi Allahyari S, Kamaliyan Z, Kadkhoda S. Circular RNA hsa_circ_0044234 as distinct molecular signature of triple negative breast cancer: a potential regulator of GATA3. Cancer Cell Int 2021; 21:312. doi: 10.1186/s12935-021-02015-6 [Crossref] [ Google Scholar]
-
Wang L, Li X, Wang D, Feng H, Ma F, Cao J, et al. Identification of circular RNAs hsa-circ-000696 as a novel biomarker for breast cancer. Research Square 2019. doi: 10.21203/rs.2.13471/v1.
- Liu J, Peng X, Liu Y, Hao R, Zhao R, Zhang L. The Diagnostic Value of Serum Exosomal Has_circ_0000615 for Breast Cancer Patients. Int J Gen Med 2021; 14:4545-54. doi: 10.2147/ijgm.S319801 [Crossref] [ Google Scholar]
- Yin WB, Yan MG, Fang X, Guo JJ, Xiong W, Zhang RP. Circulating circular RNA hsa_circ_0001785 acts as a diagnostic biomarker for breast cancer detection. Clin Chim Acta 2018; 487:363-8. doi: 10.1016/j.cca.2017.10.011 [Crossref] [ Google Scholar]
- Magalhães L, Ribeiro-Dos-Santos AM, Cruz RL, Nakamura KDM, Brianese R, Burbano R. Triple-Negative Breast Cancer circRNAome Reveals Hsa_circ_0072309 as a Potential Risk Biomarker. Cancers (Basel) 2022; 14:3280. doi: 10.3390/cancers14133280 [Crossref] [ Google Scholar]
- Li LX, Hao Y, Dong L, Qiao ZQ, Yang SC, Chen YD. Circular RNAs as Biomarkers in Breast Cancer Diagnosis, Prognosis, Molecular Types, Metastasis and Drug Resistance. Technol Cancer Res Treat 2025; 24:15330338251328500. doi: 10.1177/15330338251328500 [Crossref] [ Google Scholar]
- Chen B, Wei W, Huang X, Xie X, Kong Y, Dai D. circEPSTI1 as a Prognostic Marker and Mediator of Triple-Negative Breast Cancer Progression. Theranostics 2018; 8:4003-15. doi: 10.7150/thno.24106 [Crossref] [ Google Scholar]
- Li Y, Li H. Circular RNA VRK1 correlates with favourable prognosis, inhibits cell proliferation but promotes apoptosis in breast cancer. J Clin Lab Anal 2020; 34:e22980. doi: 10.1002/jcla.22980 [Crossref] [ Google Scholar]
- Xu J, Chen X, Sun Y, Shi Y, Teng F, Lv M. The Regulation Network and Clinical Significance of Circular RNAs in Breast Cancer. Front Oncol 2021; 11:691317. doi: 10.3389/fonc.2021.691317 [Crossref] [ Google Scholar]
- Omid-Shafaat R, Moayeri H, Rahimi K, Menbari MN, Vahabzadeh Z, Hakhamaneshi MS. Serum Circ-FAF1/Circ-ELP3: A novel potential biomarker for breast cancer diagnosis. J Clin Lab Anal 2021; 35:e24008. doi: 10.1002/jcla.24008 [Crossref] [ Google Scholar]
- Wang S, Wang Y, Li Q, Li X, Feng X. A novel circular RNA confers trastuzumab resistance in human epidermal growth factor receptor 2-positive breast cancer through regulating ferroptosis. Environ Toxicol 2022; 37:1597-607. doi: 10.1002/tox.23509 [Crossref] [ Google Scholar]
- Wu X, Ren Y, Yao R, Zhou L, Fan R. Circular RNA circ-MMP11 Contributes to Lapatinib Resistance of Breast Cancer Cells by Regulating the miR-153-3p/ANLN Axis. Front Oncol 2021; 11:639961. doi: 10.3389/fonc.2021.639961 [Crossref] [ Google Scholar]
- Wang Q, Liang D, Shen P, Yu Y, Yan Y, You W. Hsa_circ_0092276 promotes doxorubicin resistance in breast cancer cells by regulating autophagy via miR-348/ATG7 axis. Transl Oncol 2021; 14:101045. doi: 10.1016/j.tranon.2021.101045 [Crossref] [ Google Scholar]
- Yang W, Gu J, Wang X, Wang Y, Feng M, Zhou D. Inhibition of circular RNA CDR1as increases chemosensitivity of 5-FU-resistant BC cells through up-regulating miR-7. J Cell Mol Med 2019; 23:3166-77. doi: 10.1111/jcmm.14171 [Crossref] [ Google Scholar]
- Yi J, Wang L, Hu GS, Zhang YY, Du J, Ding JC. CircPVT1 promotes ER-positive breast tumorigenesis and drug resistance by targeting ESR1 and MAVS. Embo j 2023; 42:e112408. doi: 10.15252/embj.2022112408 [Crossref] [ Google Scholar]
- Liu D, Fang L. Current research on circular RNAs and their potential clinical implications in breast cancer. Cancer Biol Med 2021; 18:635-48. doi: 10.20892/j.issn.2095-3941.2020.0275 [Crossref] [ Google Scholar]
- Chen ZG, Zhao HJ, Lin L, Liu JB, Bai JZ, Wang GS. Circular RNA CirCHIPK3 promotes cell proliferation and invasion of breast cancer by sponging miR-193a/HMGB1/PI3K/AKT axis. Thorac Cancer 2020; 11:2660-71. doi: 10.1111/1759-7714.13603 [Crossref] [ Google Scholar]
- Li Z, Li Y, Han D, Wang X, Li C, Chen T. circRNA-SFMBT2 orchestrates ERα activation to drive tamoxifen resistance in breast cancer cells. Cell Death Dis 2023; 14:482. doi: 10.1038/s41419-023-06006-5 [Crossref] [ Google Scholar]
- Zang H, Li Y, Zhang X, Huang G. Circ-RNF111 contributes to paclitaxel resistance in breast cancer by elevating E2F3 expression via miR-140-5p. Thorac Cancer 2020; 11:1891-903. doi: 10.1111/1759-7714.13475 [Crossref] [ Google Scholar]
- Luo J, Zhong H, Guo M, Xiao P, Cao R, Zhao M. CircAGFG1 Promotes Ovarian Cancer Progression Through the miR-409-3 p/ZEB1 Axis. Technol Cancer Res Treat 2024; 23:15330338241252423. doi: 10.1177/15330338241252423 [Crossref] [ Google Scholar]
- Li D, Yang R, Yang L, Wang D, Zhou X, Sun Y. circANKS1B regulates FOXM1 expression and promotes cell migration and invasion by functioning as a sponge of the miR-149 in colorectal cancer. Onco Targets Ther 2019; 12:4065-4073. doi: 10.2147/OTT.S201310 [Crossref] [ Google Scholar]
- Movassagh M, Morton SU, Hehnly C, Smith J, Doan TT, Irizarry R. mirTarRnaSeq: An R/Bioconductor Statistical Package for miRNA-mRNA Target Identification and Interaction Analysis. BMC Genomics 2022; 23:439. doi: 10.1186/s12864-022-08558-w [Crossref] [ Google Scholar]