Bioimpacts. 16:33252.
doi: 10.34172/bi.33252
Review
Long non-coding RNA PURPL in cancer and cellular regulation: Mechanistic insights and therapeutic potential
Nazila Valatabar Conceptualization, Data curation, Visualization, Writing – original draft, Writing – review & editing, 1 
Reza Safaralizadeh Conceptualization, Validation, Writing – review & editing, 1, *
Amir Ali Mokhtarzadeh Conceptualization, Validation, Writing – review & editing, 2, *
Mohammad Ali Hosseinpour Feizi Validation, Writing – review & editing, 1
Author information:
1Department of Animal Biology, Faculty of Natural Science, University of Tabriz, Tabriz, Iran
2Immunology Research Center, Tabriz University of Medical Sciences, Tabriz, Iran
Abstract
Long non-coding RNAs (lncRNAs) constitute a large fraction of the mammalian transcriptome and are increasingly recognized as critical regulators of gene expression and cellular homeostasis. Dysregulation of lncRNAs has been strongly implicated in cancer initiation and progression. PURPL (p53-upregulated regulator of p53 levels) is an emerging oncogenic lncRNA that plays a pivotal role in modulating multiple cancer-related cellular pathways, including the p53 signaling pathway, autophagy, miRNA-mediated gene regulation, genome stability, cellular senescence, and metastasis. Mechanistically, PURPL orchestrates signaling networks and epigenetic regulation via diverse molecular modes, functioning as a competing endogenous RNA (ceRNA), molecular scaffold, and guide, thereby influencing gene expression and cellular homeostasis. Functional studies reveal that suppression of PURPL enhances chemosensitivity, inhibits tumor cell proliferation and migration, and induces apoptosis across multiple cancer types. This review systematically summarizes the molecular mechanisms underlying PURPL-mediated regulation and highlights its multifaceted roles in cancer biology, thereby providing an integrative view of its functions in tumorigenesis and cellular regulation.
Graphical Abstract
Keywords: LncRNA, PURPL, p53 Signaling pathway, Cancer progression, Autophagy, Competing endogenous RNA, Metastasis, Chemosensitivity
Copyright and License Information
© 2026 The Author(s).
This work is published by BioImpacts as an open access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (
http://creativecommons.org/licenses/by-nc/4.0/). Non-commercial uses of the work are permitted, provided the original work is properly cited.
Funding Statement
The authors are grateful for financial support from the Immunology Research Center, Tabriz University of Medical Science (Grant number: 78109).
Introduction
lncRNAs are RNA molecules longer than 200 nucleotides and represent a substantial portion of the ~98% of mammalian genomic transcripts that do not code for proteins.1-3 Although they lack an open reading frame and do not encode proteins, lncRNAs share several features with messenger RNAs (mRNAs), including transcription by RNA polymerase II, the presence of a 5' methylated cap, and a 3' polyadenylated tail.2,4-6 They can be grouped into four main types depending on how they’re positioned relative to nearby protein-coding genes.2,7 Sense lncRNAs overlap with one or more exons on the same DNA strand. On the other hand, antisense lncRNAs overlap exons but on the opposite strand. Intronic lncRNAs originate from introns within other gene transcripts, while intergenic lncRNAs are found in the regions between two genes on the same strand.2,7 lncRNAs engage in extensive molecular interactions that enable them to regulate diverse intracellular processes.5-8 Primarily, they function as signaling molecules, either independently or in association with other proteins such as transcription factors, to modulate the transcription of downstream target genes.9 Additionally, lncRNAs can act as molecular decoys by directly binding to functional proteins, including those involved in chromosome architecture or transcriptional regulation, thereby inhibiting their activity.9
Moreover, lncRNAs have the capacity to bind microRNAs (miRNAs), acting as molecular sponges that sequester miRNAs and prevent their interaction with target mRNAs, consequently leading to upregulation of the respective target genes.9 Furthermore, lncRNAs serve as guide molecules, facilitating the recruitment of specific proteins such as transcription factors to precise genomic loci, thereby influencing gene transcription.9,10 They also function as scaffolds, providing a platform for the assembly of macromolecular complexes, including proteins, to orchestrate downstream signaling pathways.9,10 In addition to these roles, lncRNAs are critically involved in epigenetic regulation.10 Therefore, due to these critical functions, any disruption in the regulatory mechanisms of lncRNAs can disturb cellular homeostasis and contribute to the development of diseases like malignancies.11 Since first being highlighted in 2017, PURPL lncRNA “p53 upregulated regulator of p53 levels, LINC01021”, as a member of the lncRNA family, has been a subject of research interest due to its regulatory functions in cancer-related processes.12,13 PURPL is an intergenic lncRNA positioned between protein-coding genes. It has been identified as a key regulator of the p53 protein, which is essential for maintaining genomic integrity. Additionally, extensive research has demonstrated that PURPL is involved in other critical cellular processes, including autophagy and the regulation of miRNAs function. Dysregulated expression profile and altered activity of PURPL have been documented in numerous malignancies, highlighting its oncogenic role. This review aims to comprehensively summarize the current understanding of PURPL, with a particular focus on its regulatory mechanisms and its functional roles in cancer biology.
Molecular features and expression profile of PURPL
This intergenic long non-coding RNA is located on the plus strand of chromosome 5 at position p14.1 and encompasses a length of 24,110 base pairs. Alternative splicing events give rise to multiple PURPL transcript variants, which differ in exon content and composition; the full-length variant contains five exons. Thus, it appears that each variant potentially contributes to diverse functional roles in cellular processes. Multiple studies have indicated that PURPL expression varies between normal and cancerous tissues. In healthy cells, PURPL is typically expressed at low to moderate levels (e.g., 0.5–5 TPM, based on TCGA RNA sequencing data), contributing to normal cellular homeostasis and function. However, numerous reports have documented aberrant overexpression or dysregulation of PURPL in various cancers, including colorectal, liver, melanoma, Gastric, and breast cancers, where its expression can increase significantly. To better understand the molecular basis of these observations, the following section focuses on the regulatory mechanisms that govern PURPL expression and function.
PURPL function in relation to p53
PURPL as a modulator of p53 signaling in cancer regulation
The tumor suppressor p53 is essential for preserving genomic stability by pausing the cell cycle following DNA damage, facilitating repair processes, and triggering apoptosis when damage is beyond repair to eliminate potentially harmful cells.14,15 Under normal conditions without cellular stress, p53 remains inactive due to continuous ubiquitination by its negative regulator MDM2, which targets it for proteasomal degradation.14-16 However, in response to stress or DNA damage, the protein Myb-binding protein 1A (MYBBP1A) relocates from the nucleolus to the nucleoplasm and facilitates p53 tetramerization.17 This conformational change reduces p53’s binding affinity to MDM2, thereby stabilizing the protein and enhancing its transcriptional activity in stress response pathways. Evidence also suggests that p53 may regulate PURPL expression, which itself contributes to cell survival in response to DNA damage. Furthermore, studies have shown that PURPL interacts with MYBBP1A, disrupting the formation of the MYBBP1A–p53 complex and thereby inhibiting p53 activation. This highlights PURPL’s regulatory role in the functional pathway of p53 (Fig. 1).12
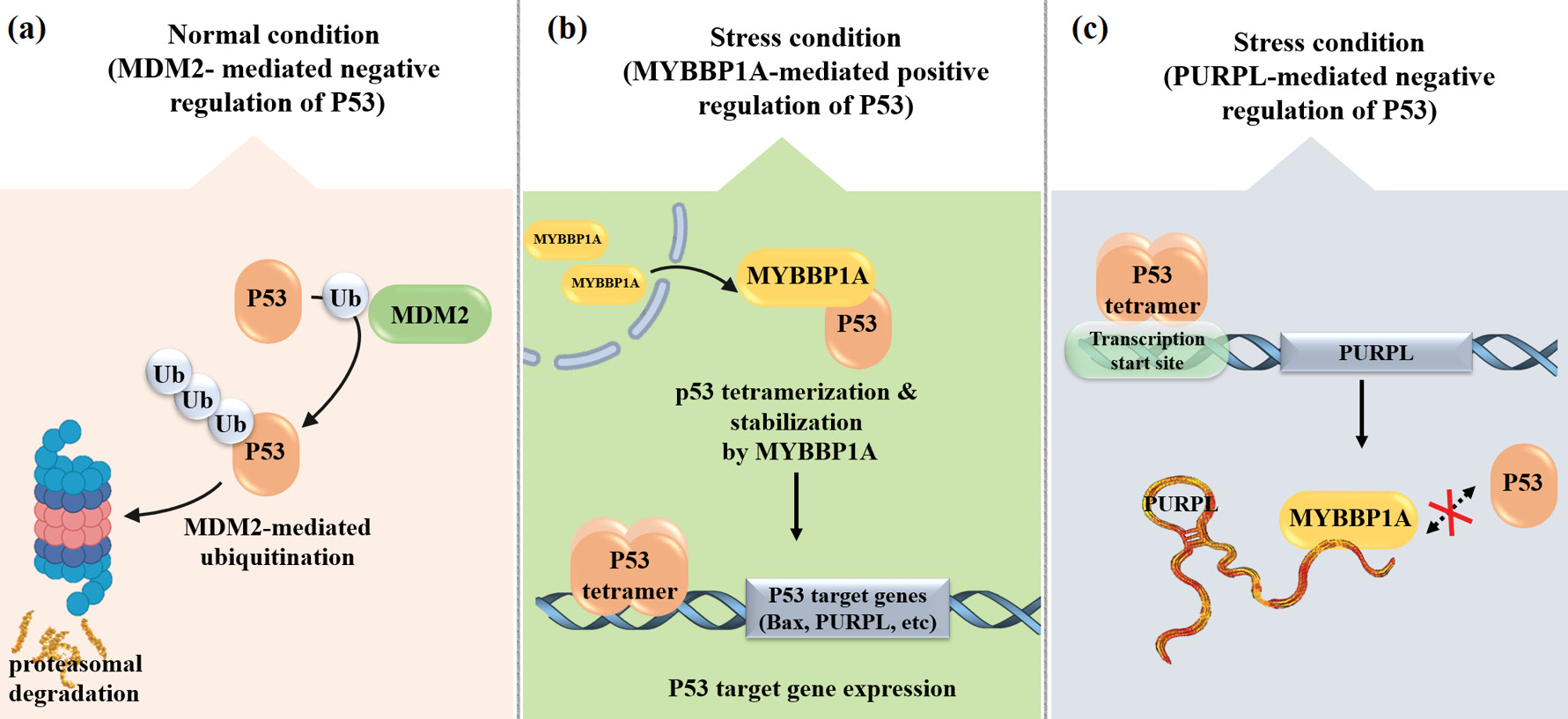
Fig. 1.
Schematic representation of (a, b) the negative regulation of p53 by MDM2 under normal conditions and its positive regulation by MYBBP1A under stress conditions, and (c) the negative regulation of p53 by the lncRNA PURPL.
.
Schematic representation of (a, b) the negative regulation of p53 by MDM2 under normal conditions and its positive regulation by MYBBP1A under stress conditions, and (c) the negative regulation of p53 by the lncRNA PURPL.
PURPL as a modulator of p53 signaling in cellular senescence
One stable and irreversible type of cell cycle stoppage is cellular senescence. Many cellular stresses, such as oxidative stress, telomere shortening, DNA damage, and metabolic changes, can cause this. Crucially, the two mechanisms that control cellular senescence—the p53/p21 and p16^INK4a/Rb pathways—induce and sustain cellular senescence by activating the DNA damage response (DDR) and causing the cell cycle to end. Significant molecular and structural alterations in the senescent cell, such as chromatin organization remodeling, modifications to transcriptional programming, and adjustments to metabolic activity, are hallmarks of senescence. Additionally, they develop a Senescence-Associated Secretory Phenotype (SASP), secreting growth factors, proteases, and pro-inflammatory cytokines that alter the tissue milieu, cause chronic inflammation, and may eventually encourage cancer or tissue malfunction.18
In a study, it was shown that PURPL is involved not only in oncogenic processes but also in cellular aging. PURPL has been linked to both cellular aging and carcinogenic processes, according to a study.19 In replicative senescence models, aged VSMCs displayed morphological alterations, cell-cycle arrest in G1 phase, activation of the DNA damage response (DDR), and traditional senescence markers such as elevated p53, p21, p16, and SA-β-gal activity.19 Concurrently, there was a marked upregulation of PURPL expression, indicating its role in the senescence program. According to these results, PURPL is essential for controlling cell cycle arrest and stress adaptation during vascular aging.19 Mechanistically, PURPL modulates the p53 network. So, it has been suggested that PURPL elevation may act as a positive feedback modulator in VSMCs, senescent to fine-regulating p53 activity, preventing early or extreme senescence. The siRNA-mediated knockdown experiments and correlation analysis revealed that downregulation of PURPL impairs the p53-dependent transcriptional pathway, thus disrupting the equilibrium between cell survival and irreversible growth arrest.19
The upregulation of PURPL during senescence most likely reflects a protective adaptive response that helps to stabilize cells under stress and DNA damage. On the other hand, continuous overexpression may cause vascular dysfunction by sustaining senescent phenotypes, boosting inflammatory signaling via SASP molecules, and modifying tissue remodeling pathways.19 Taken together, evidence indicates that PURPL functions as a molecular regulator for p53 signaling in cellular aging by adjusting the beneficial and adverse effects of senescence. This lncRNA thus has a dual role, promoting oncogenesis under conditions of oncogenic stress and maintaining homeostasis in vascular senescence, which highlights the complex role of lncRNA in regulating cell networks in both physiological and pathological contexts.19
PURPL and genomic integrity
CIN is a common property of many tumors and is characterized by aberrant chromosome segregation during mitosis that leads to cells with aneuploidy or structural chromosomal rearrangements.20 CIN creates genetic heterogeneity within tumor cells, with some cells becoming fitter and able to survive adverse conditions (including anticancer therapies), while others are compromised or die from the genomic stress caused by instability.20 CIN can both drive and limit tumor growth, survival, and progression, depending on the genetic diversity and microenvironmental responses; recent evidence indicates that CIN and aneuploidy can also trigger inflammatory responses in the tumor microenvironment that initially may serve as antitumor mechanisms but can later lead to a favorable niche for cancer cell survival and metastasis.21 Together, CIN is not only a secondary characteristic of cancer, but also a major cause of tumor progression, metastasis, and therapy resistance.20,21 In this regard, lncRNA PURPL has been identified as an important regulator of chromosomal stability. Disruption of its expression can markedly disturb the balance between genome maintenance and genomic instability. Notably, previous studies have reported significant upregulation of PURPL in various models of chromosomal instability (CIN) and aneuploidy. This increase has been observed in cells treated with CIN-inducing agents, including reversine, cytochalasin B, and ZM447439. Importantly, PURPL upregulation in these contexts is largely dependent on functional p53 status.22 When p53 is activated in response to genomic stress, PURPL is upregulated as a part of the cellular response to genomic stress and helps to regulate the balance of the p53 pathway by regulating MDM2, a negative regulator of p53.22 However, loss or knockdown of PURPL in normal cells results in CIN phenotypes, nuclear morphological alterations, increased sensitivity to aneuploidy-inducing agents, and defective chromosome segregation, suggesting that precise regulation of PURPL expression is essential: overexpression may cause genomically unstable cells to survive via p53 over-inhibition, while reduced expression can lead to loss of control over chromosomal stability.22 Therefore, PURPL functions as a node within the p53–MDM2 axis that plays a role in maintaining or disrupting genomic stability.22
Overall, PURPL functions as a versatile and context-dependent regulator, intricately modulating p53 signaling, cellular senescence, and genomic integrity. In cancer cells, it often dampens p53 activity to promote survival and proliferation, whereas in normal or aging cells, it may support adaptive senescence and maintain cellular homeostasis. The dual and sometimes opposing roles of PURPL underscore its context-specific effects, which are influenced by p53 status, cell type, and experimental conditions. These observations highlight critical gaps in our understanding and emphasize the need for further studies to delineate its oncogenic versus homeostatic functions, as well as its potential as a therapeutic target across different biological and pathological contexts.
While the interaction between PURPL and the p53 signaling pathway underscores its role in genomic stability and stress responses, PURPL also exerts broad regulatory effects through post-transcriptional mechanisms, particularly via its interactions with microRNAs, which are discussed in the next section.
Regulatory network of PURPL and miRNAs
Post-transcriptional regulation of gene expression is a fundamental function of lncRNAs, many of which exert their regulatory effects through interactions with miRNAs. In this context, lncRNAs can function as ceRNAs by harboring binding sites for specific miRNAs and sequestering them via a sponging mechanism, thereby reducing miRNA availability for binding to the 3′ untranslated regions (3′ UTRs) of target mRNAs.9,23,24 This interaction limits miRNA-mediated translational repression or mRNA degradation and ultimately modulates the expression of downstream genes.25 As key post-transcriptional regulators, miRNAs play essential roles in controlling cellular processes such as proliferation, apoptosis, migration, and differentiation; consequently, disruption of lncRNA–miRNA regulatory networks is frequently associated with tumorigenesis and disease progression.26-28
Within this regulatory framework, accumulating experimental evidence indicates that PURPL engages in specific miRNA-centered regulatory axes, and several representative studies highlighting the functional consequences of PURPL–miRNA interactions in cancer are discussed below.
PURPL/miR-363-3p/ADAM10 axis
miR-363-3p has been linked with numerous types of cancer, including ovarian serous cystadenocarcinoma, liver, stomach, glioma, prostate, lung, osteosarcoma, and breast cancers. It targets multiple pathways, suppressing cell growth and migration via SOX4 and NEDD9 targeting and enhancing proliferation via XIST interaction and MDM2 upregulation.29 In ovarian serous cystadenocarcinoma, bioinformatics analysis and experimental data both indicated PURPL's involvement in the regulation of the miR-363-3p/ADAM10 pathway.30 Specifically, miR-363-3p was determined to target ADAM10 directly, a transmembrane metalloproteinase that sheds ligands, receptors, and cell adhesion molecules. ADAM10 cleavage is often a precursor to regulated intramembrane proteolysis (RIP). A significant substrate is the Notch receptor: ADAM10 cuts its extracellular domain following ligand binding, with γ-secretase cutting of the remaining fragment, releasing the Notch intracellular domain (NICD), which translocates to the nucleus to regulate genes implicated in cell growth, differentiation, and survival. Dysfunctional Notch is strongly associated with tumorigenesis.31,32 PURPL acts as a molecular sponge for miR-363-3p, relieving ADAM10 from miRNA-mediated repression.30 This upregulates ADAM10 and Notch pathway activation expression, promoting proliferation, migration, and invasion. The PURPL/miR-363-3p/ADAM10 axis is thus a key regulatory pathway in ovarian serous cystadenocarcinoma and has been proposed to be a therapeutic target or prognostic biomarker.30
The TAM–PURPL/miR-363/PDZD2 signaling axis and immune crosstalk
Recent research has shown that the lncRNA PURPL/miR-363/PDZD2 axis is a key promoter of osteosarcoma (OS) progression and metastasis. Tumor-associated macrophages (TAMs), as one of the most significant components of the tumor microenvironment (TME), have been proven to play a critical role in OS development. TAMs, predominantly recruited from circulating monocytes, infiltrate into the tumor tissues and acquire an M2-like phenotype with immunosuppressive, pro-tumorigenic properties.33-36 These macrophages secrete cytokines, growth factors, and extracellular vesicles that promote tumor growth, angiogenesis, and epithelial–mesenchymal transition (EMT), and suppress antitumor immunity.33-37 Recent findings demonstrate that TAMs elevate the levels of lncRNA PURPL in OS cells, suggesting PURPL as a downstream mediator of macrophage-derived signals. Elevated PURPL levels then act as an upstream regulator of miR-363, a tumor suppressor miRNA. It was reported that under normal circumstances, miR-363 represses the expression of PDZD2 (PDZ domain containing 2), a gene implicated in cell proliferation, migration, and invasion. In TAM-stimulated OS cells, though, PURPL binds and sequesters miR-363, relieving PDZD2 from miRNA-mediated inhibition.37
As a result, PDZD2 overexpression sets off a chain of oncogenic events, including elevated cell proliferation, motility, invasiveness, and EMT, all of which favor tumor aggressiveness and metastatic dissemination. Functional investigations indicated that depletion of PURPL or PDZD2 reverses these malignant phenotypes and compromises TAM-induced tumor growth, whereas miR-363 replenishment exerts tumor suppressive roles.37 Collectively, these findings establish that the TAM–PURPL/miR-363/PDZD2 signaling axis represents a critical link between immune cell–derived signals and noncoding RNA–mediated gene expression in OS (Fig. 2).37
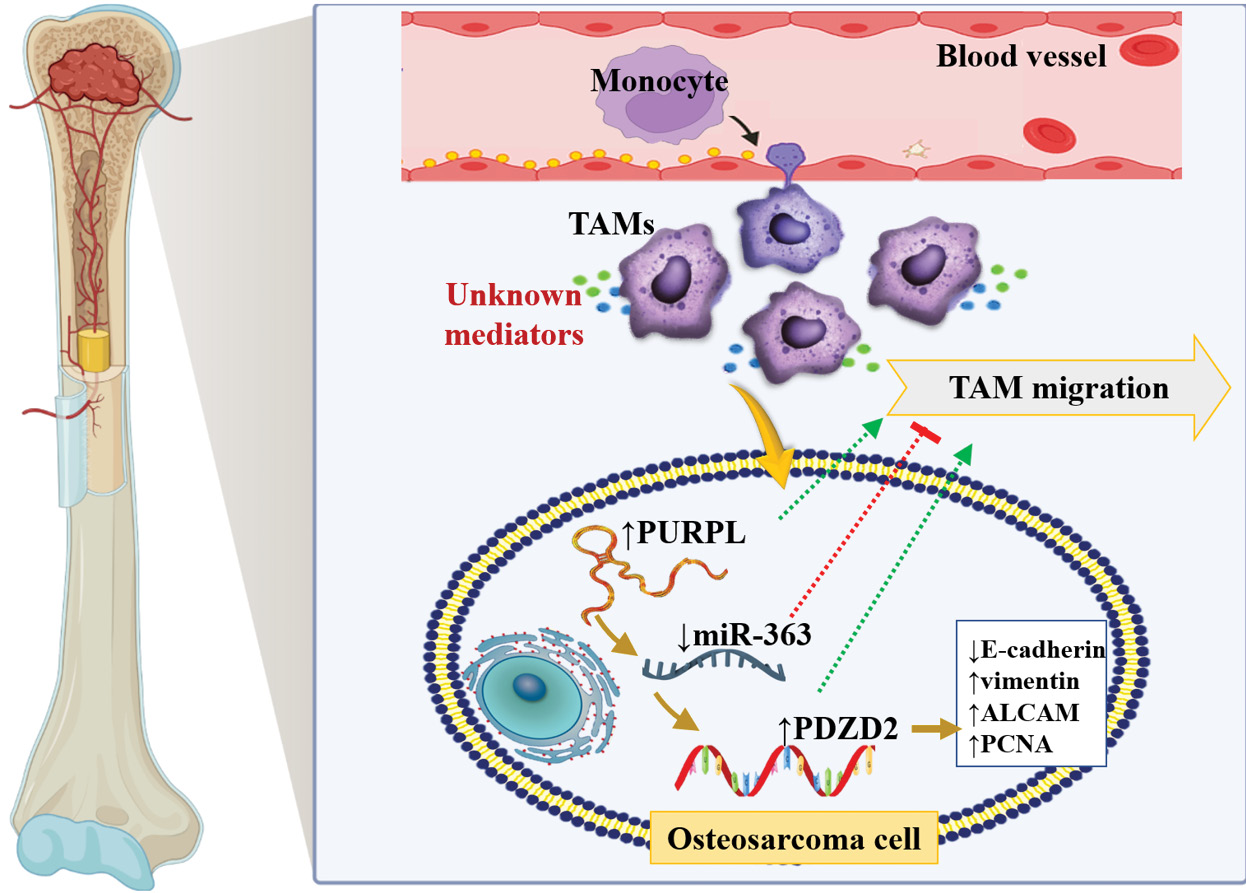
Fig. 2.
The interaction between TAMs and the PURPL/miR-363/PDZD2 axis contributes to OS progression. TAMs may enhance OS development by modulating PURPL/miR-363/PDZD2 signaling, while this signaling axis can, in turn, provide feedback regulation that influences TAM migration.
.
The interaction between TAMs and the PURPL/miR-363/PDZD2 axis contributes to OS progression. TAMs may enhance OS development by modulating PURPL/miR-363/PDZD2 signaling, while this signaling axis can, in turn, provide feedback regulation that influences TAM migration.
PURPL and MiR-338-3p axis
MiR-338-3p is involved in multiple cellular processes, comprising proliferation, apoptosis, migration, and invasion, and is implicated in tumor initiation and progression in many human cancers, including colorectal, gastric, breast, lung, and epithelial ovarian carcinomas.38 For example, in ovarian cancer, miR-338-3p downregulation has been linked to increased tumor cell growth and metastasis, which could be due to the regulation of cell proliferation and EMT by MACC1, Met, and downstream Wnt/β-catenin and MEK/ERK signaling pathways,39 and it has been found that downregulation of miR-338-3p is correlated with chemoresistance, whereas overexpression sensitizes cancer cells to cisplatin, sorafenib, doxorubicin, vinblastine, and 5-FU.38 In addition, recent evidence suggests that lncRNAs, especially PURPL, may act as upstream regulators of miR-338-3p, suppressing its expression and thereby driving the progression of ovarian cancer. Furthermore, the combination of downregulated miR-338-3p and upregulated PURPL was significantly associated with poor RFS and OS in patients with epithelial ovarian cancer, implying that dysregulation of this axis may serve as a potential prognostic biomarker and therapeutic target.39
PURPL-mediated oncogenic regulation via the miR‑137/ZBTB7A/PI3K‑NF‑κB axis
Zhonghua Cheng et al. identified PURPL as the critical oncogenic factor in gastric cancer. The analyses of gastric cancer tissues and cell lines revealed overexpression of PURPL compared to normal controls. Functional experiments demonstrated that high levels of PURPL enhance colony formation, cellular proliferation, migration, and invasion and inhibit apoptosis in tumor cells.40
It was shown that PURPL exerted oncogenic functions in different types of cancers through the regulation of important signaling pathways. The mechanistic studies indicated that PURPL modulates the miR-137/ZBTB7A/PI3K-NF-κB signaling axis, which promotes survival, proliferation, and invasive behavior in tumor cells (Fig. 3).40
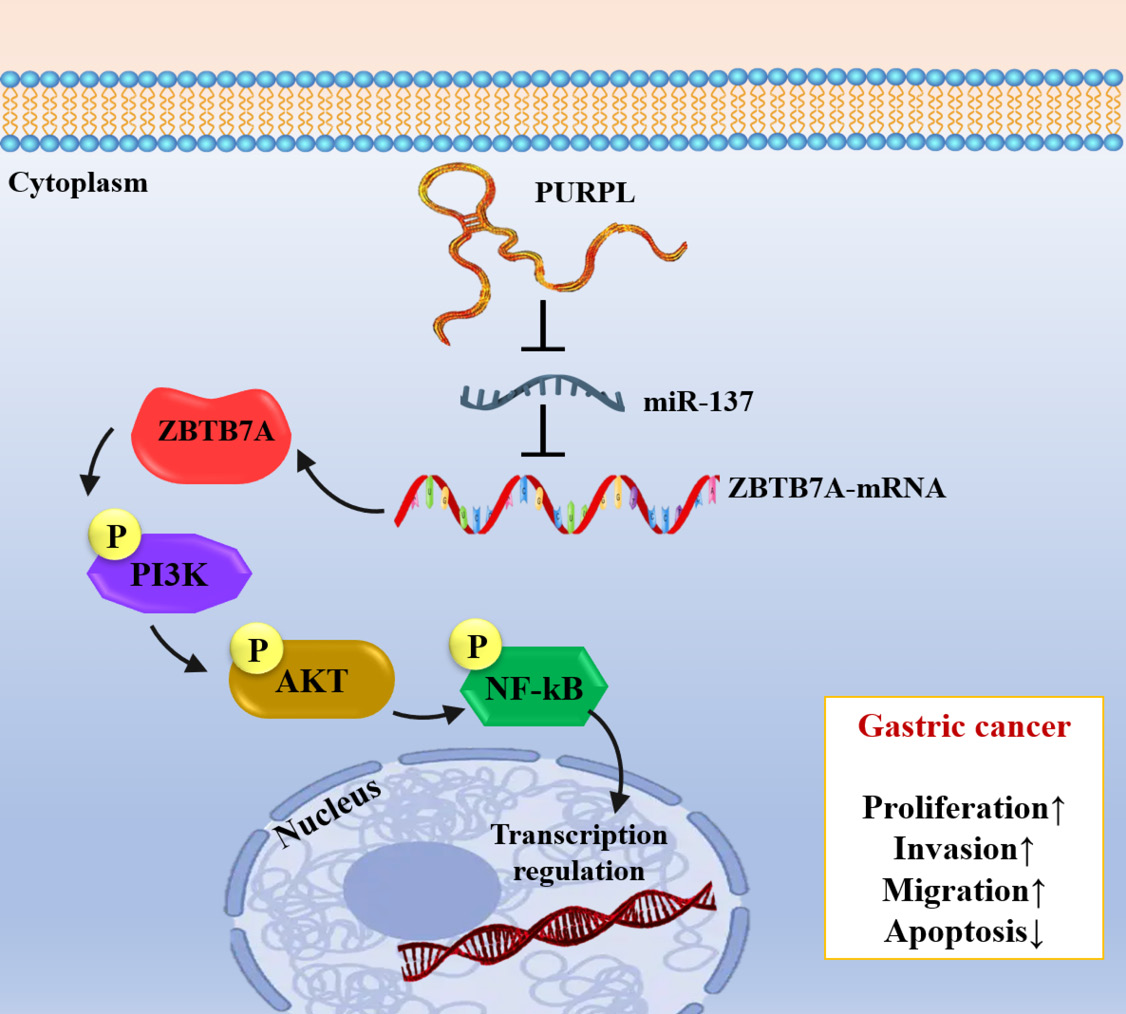
Fig. 3.
Schematic illustration of PURPL-miR-137-ZBTB7A/PI3K/AKT/NF-κB axis in gastric cancer development.
.
Schematic illustration of PURPL-miR-137-ZBTB7A/PI3K/AKT/NF-κB axis in gastric cancer development.
Numerous studies have shown that the downregulation of miR-137, a tumor-suppressive miRNA, is correlated with the progression of various cancers, including lung, colorectal, glioma, and gastric cancers.41 In this situation, elevated PURPL inhibits miR-137 expression, which raises ZBTB7A activity and activates downstream PI3K/AKT/NF-κB signaling, all of which support carcinogenic traits.40 Additionally, in vivo research confirmed that overexpression of PURPL stimulates the growth of animal tumors, while pharmacological inhibition of NF-κB by JSH-23 or PI3K by LY294002 considerably reduced these carcinogenic effects, decreasing the invasive and proliferative characteristics of tumor tissues.40 These results suggest that the PURPL-miR-137-ZBTB7A-PI3K/AKT/NF-κB axis may be a therapeutic target in cancer treatment.40
Beyond its role in miRNA-mediated regulation, PURPL also exerts profound effects on cancer progression through epigenetic regulatory mechanisms, which are discussed in the following section.
PURPL-mediated epigenetic regulation of cancer progression
Numerous cancers are known to arise from aberrant epigenetic events, including aberrant DNA methylation, altered histone modifications, and disrupted chromatin architecture.42 For example, in a study of colorectal cancer (CRC), lncRNA-PURPL was shown to epigenetically regulate tumorigenesis through direct binding to the m6A reader protein IMP2 (IGF2BP2) to increase its stability and activity.42 IMP2 then binds to m6A-modified regions on oncogenic transcripts as an III N6-methyladenosine (m6A) reader, resulting in enhanced post-transcriptional stability and translation of the target mRNAs, including MSX1 and JARID2, which were found to be important regulatory genes in this pathway (Fig. 4, Table 1).42 MSX1, a transcription factor that plays a critical role in embryonic development and cell fate determination, has been reported to activate the FGF–MAPK signaling pathway and to downregulate the tumor suppressor gene FBXW7, which stabilizes oncogenic proteins like c-MYC and MCL1.43 These events ultimately lead to increased proliferation, reduced apoptosis, and increased drug resistance in cancer cells, indicating the importance of the MSX1–FBXW7 axis as a molecular mechanism in tumor progression.43,44 On the other hand, the non-catalytic regulatory subunit JARID2 of the Polycomb Repressive Complex 2 (PRC2) directs the histone methylation of H3K27me3 on chromatin and epigenetically silences tumor suppressor genes such as PTEN to regulate the cell cycle progression, differentiation, survival, and migration of tumor cells.45 In summary, PURPL plays a pivotal role in the PURPL–IMP2–MSX1/JARID2 axis that stabilizes oncogenic mRNAs through coordinated RNA- and chromatin-based epigenetic regulation to promote cancer progression.42
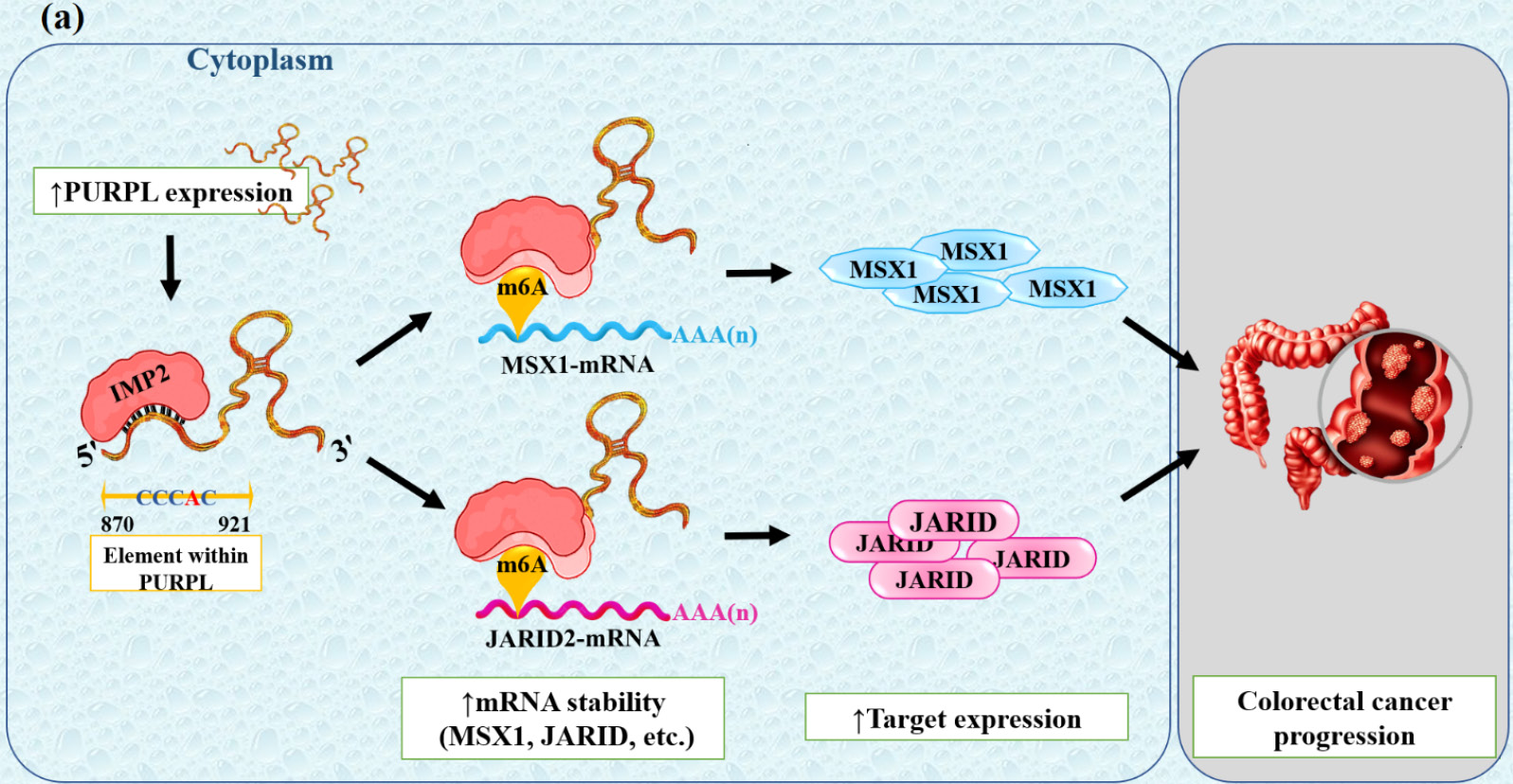
Fig. 4.
(a) The model illustration of the PURPL–IMP2–MSX1/JARID2 signaling cascade regulations and functions in CRC cells. Expression of PURPL as well as IMP2 was increased in CRC. IMP2 binds directly to the 'CCCAC' sequence of PURPL and, through m6A-dependent post-transcriptional regulation, increases the stability of its downstream transcripts MSX1 and JARID2, thereby contributing to CRC onset and progression.
.
(a) The model illustration of the PURPL–IMP2–MSX1/JARID2 signaling cascade regulations and functions in CRC cells. Expression of PURPL as well as IMP2 was increased in CRC. IMP2 binds directly to the 'CCCAC' sequence of PURPL and, through m6A-dependent post-transcriptional regulation, increases the stability of its downstream transcripts MSX1 and JARID2, thereby contributing to CRC onset and progression.
Table 1.
Functional Roles and Molecular Mechanisms of lncRNA PURPL in Cellular Processes and Disease
|
Function/Axis
|
Molecular features and mechanisms
|
Pathways and mechanisms
|
Cancers and models
|
Biological and clinical outcomes
|
Ref.
|
| p53 Regulation |
Inhibition of p53 activity via interaction with MYBBP1A |
Inhibits MYBBP1A–p53 complex formation → reduced p53 activation |
HCC, CRC, cellular models |
Increased cell survival and proliferation, decreased apoptosis |
12
|
| MiRNA regulation (ceRNA) |
Acts as a molecular sponge for specific miRNAs |
PURPL/miR-363-3p/ADAM10, PURPL/miR-363/PDZD2, PURPL/miR-338-3p, PURPL/miR-137/ZBTB7A axes
|
Ovarian, Osteosarcoma (OS), Gastric, Colorectal |
Promotes proliferation, migration, invasion, drug resistance, and poor prognosis |
30,37,39,40
|
| Epigenetic regulation |
Stabilizes oncogenic mRNAs and suppresses tumor suppressor genes |
PURPL–IMP2–MSX1/JARID2 axis
|
CRC |
Increased proliferation, decreased apoptosis, enhanced chemoresistance |
42
|
| Chromosomal instability (CIN) |
Maintains genomic balance and prevents chromosomal instability |
↑PURPL → survival of genomically unstable cells
|
Cell models, CIN-inducing drug models |
Protects cancer cells from genomic stress and therapy |
22
|
| Autophagy regulation |
Inhibits autophagy via mTOR and ULK1 |
↑mTOR-mediated ULK1 phosphorylation → autophagy inhibition |
Melanoma |
Enhanced tumor cell survival and proliferation, reduced autophagic cell death |
46
|
| Tumor growth and metastasis |
CDK2–CDX2–KISS1 axis modulation |
↑CDK2 → CDX2 phosphorylation → ↓KISS1 |
Gastric cancer |
Increased migration, invasion, EMT, and metastasis |
1
|
| Cellular senescence |
Positive modulation of p53 in aged cells |
Balances cell survival and cell cycle arrest |
VSMCs, aging models |
Maintains homeostasis and stress response, may contribute to vascular dysfunction |
19
|
| Chemosensitivity |
Knockdown enhances apoptosis and reduces proliferation |
Modulates p53-dependent downstream programs |
HCC, CRC |
Increased sensitivity to chemotherapeutic agents such as doxorubicin and 5-FU |
47,48
|
Notably, PURPL’s impact on cancer is not limited to epigenetic regulation; it also affects autophagy and metastasis, as described in the following sections.
PURPL and autophagy regulation
Autophagy is a key homeostatic process that degrades and recycles damaged organelles and aggregated proteins to maintain cell survival and genomic stability, and it is critical for intracellular recycling in response to metabolic stress or nutrient deprivation under normal physiological conditions.49 Autophagy has a dual role in cancer, acting as a protective tumor-suppressive mechanism during the early stages of tumorigenesis by removing damaged cells and reducing oxidative stress, while cancer cells exploit autophagy as a survival mechanism in late-stage cancer under conditions of hypoxia, nutrient limitation, and therapeutic stress.49 In this context, autophagy promotes tumor growth, metastasis, and drug resistance. The key signaling pathways that regulate autophagy are mainly the mechanistic target of rapamycin (mTOR), AMPK, and p53.50 In cancer cells, mTOR inhibition can either lead to cell death and inhibition of tumor growth or, conversely, can stimulate cell survival in hostile microenvironments.
AMPK acts as an energy sensor, becoming activated when ATP levels are low. It inhibits mTOR activity and directly activates genes related to autophagy. This process helps remove damaged cells and maintain cellular balance in healthy cells. However, in cancer cells, AMPK activation may provide energy and increase stress resistance, especially in low-oxygen conditions or during treatment with anticancer therapies.50 The outcome of autophagy, as controlled by AMPK and related pathways, is determined in large part by the tumor suppressor p53: in cells with functional p53, autophagy induced by AMPK and related pathways eliminates damaged or genetically unstable cells with tumor-suppressive effects, whereas in cancer cells with inactivated or mutant p53, the control of autophagy is lost, allowing cancer cells to survive through energy and metabolic homeostasis.50
A recent study showed that the PURPL regulates autophagy in cancer and is highly upregulated in melanoma cells and cutaneous melanoma tumor samples.46 PURPL overexpression is associated with increased cell proliferation, colony formation, migration, and invasion, which is consistent with its function as an oncogene.46 Mechanistically, PURPL directly binds to mTOR and ULK1, and high PURPL levels increase mTOR-mediated ULK1 phosphorylation at Ser757, which inhibits autophagy and prevents autophagic cell death, promoting tumor cell survival under metabolic or therapeutic stress.46 On the other hand, PURPL knockdown or suppression activates the AMPK pathway, resulting in increased ULK1 phosphorylation at Ser555 and Ser317, which enhances autophagy and autophagy-mediated cell death, significantly reducing melanoma cell growth.46 This process severely inhibits the growth of melanoma cells. These findings were validated in vivo using mouse models in which PURPL knockdown dramatically decreased tumor volume and suppressed tumor aggressiveness, indicating that PURPL knockdown can induce autophagic cell death and prevent tumor progression (Fig. 5, Table 1).46
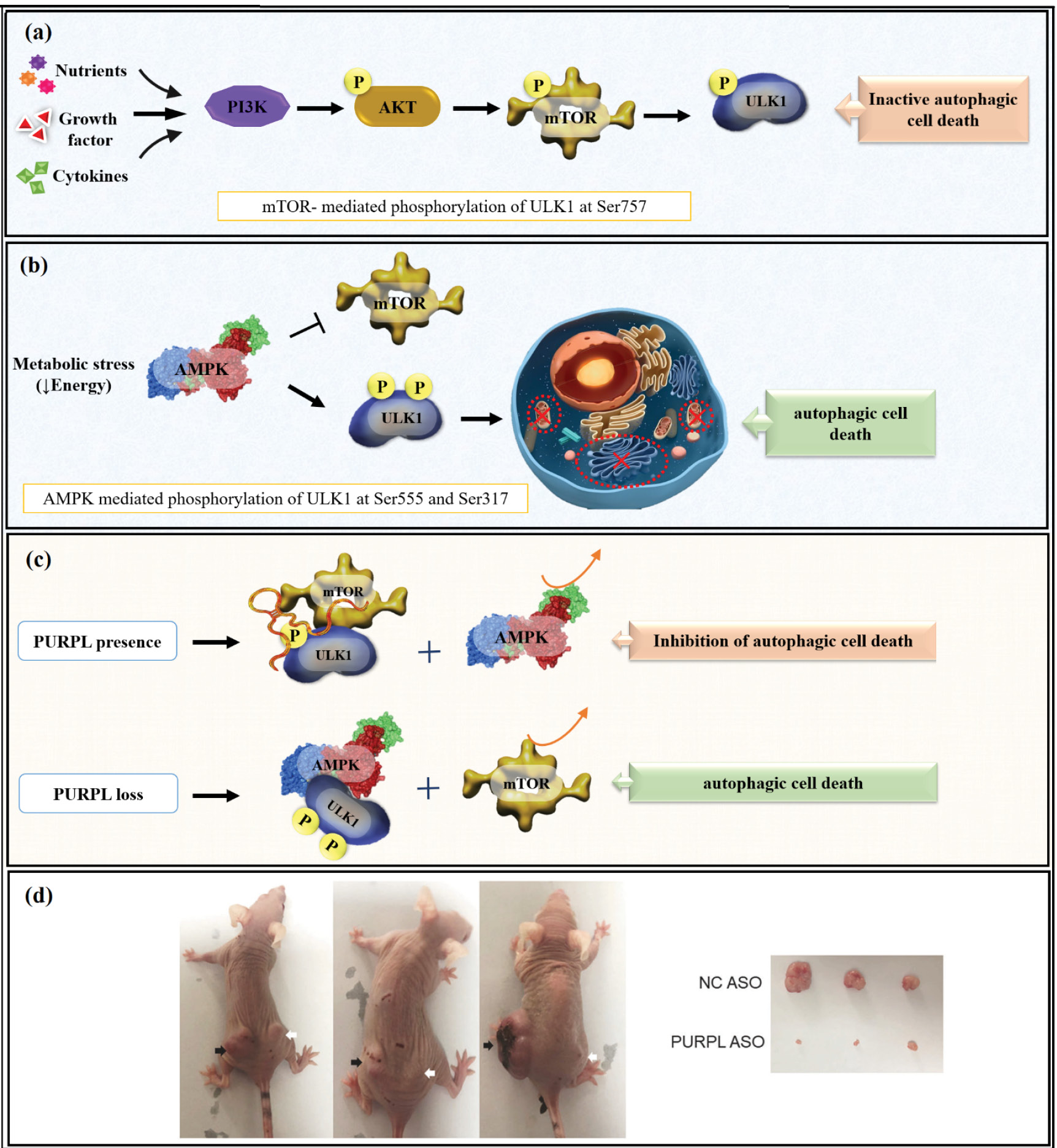
Fig. 5.
(a, b) Showing negative and positive regulation of autophagic cell death through mTOR-mediated and AMPK-mediated phosphorylation, and (c) inhibitory role ofPURPL in autophagic cell death by interaction with mTOR and ULK. (d) Representative imaging of xenograft tumor weight and tumor size. Knockdown of PURPL suppressed the growth rate and weight of tumors in xenografts. (Note: black arrows show the NC ASO-treated xenografts while white arrows show PURPL ASO-treated xenografts). Adapted from Han et al
46
under a Creative Commons Attribution 4.0 International License.
.
(a, b) Showing negative and positive regulation of autophagic cell death through mTOR-mediated and AMPK-mediated phosphorylation, and (c) inhibitory role ofPURPL in autophagic cell death by interaction with mTOR and ULK. (d) Representative imaging of xenograft tumor weight and tumor size. Knockdown of PURPL suppressed the growth rate and weight of tumors in xenografts. (Note: black arrows show the NC ASO-treated xenografts while white arrows show PURPL ASO-treated xenografts). Adapted from Han et al
46
under a Creative Commons Attribution 4.0 International License.
PURPL as a regulator of metastatic progression
One of the defining hallmarks of cancer is metastasis, which accounts for most cancer deaths. This multistep process involves local invasion of cancer cells into surrounding tissues, degradation of the ECM by enzymes such as matrix metalloproteinases (MMPs), intravasation into the bloodstream or lymphatic system, survival of circulatory stress and immune surveillance, extravasation into distant organs, adaptation to the new microenvironment, and formation of secondary tumors.51 Key molecular mechanisms include epithelial–mesenchymal transition (EMT), which is promoted by signaling pathways like TGF-β, Wnt/β-catenin, Notch, and PI3K/AKT, leading to cellular migration and invasion; activation of the PI3K/AKT and MAPK pathways to promote cell survival under metabolic and oxidative stress; and NF-κB signaling, which supports inflammatory responses, immune evasion, and metastatic niche establishment, all of which contribute to the survival, adaptation, and proliferation of cancer cells in secondary sites, leading to tumor progression, therapeutic resistance, and poor patient survival.52
Recent evidence has highlighted PURPL as an important oncogenic driver in gastric cancer progression.1 It has been reported that PURPL promotes tumor growth and metastasis by modulating the CDK2–CDX2–KISS1 signaling axis. Under normal conditions, the transcription factor CDX2 binds directly to the promoter of the metastasis suppressor KISS1 and induces its transcription, which limits tumor invasion and spread.1 However, PURPL upregulation enhances CDK2 (cyclin-dependent kinase 2) activity, which in turn phosphorylates CDX2 at serine residues, inactivating it and causing it to be retained in the cytoplasm and thus unable to bind to the KISS1 promoter, leading to a decrease in KISS1 expression (Fig. 6, Table 1).1 The inhibition of KISS1 prevents its repression of MMPs and metastasis-related signaling pathways, which increases cell migration, invasion, and EMT.1 On the other hand, silencing or knockdown of PURPL prevents CDK2-mediated phosphorylation of CDX2, enabling its nuclear translocation and transcriptional activity, reactivating KISS1 to suppress tumor cell motility, decrease metastatic potential, and induce apoptotic cell death.1 Knockdown of PURPL was confirmed to effectively suppress tumor growth and metastasis in functional assays in vitro (cell proliferation, colony formation, migration, and invasion) and in vivo (xenograft mouse models).1 Collectively, these findings reveal a clear oncogenic mechanism whereby PURPL acts as an upstream regulator of the CDK2–CDX2–KISS1 axis.
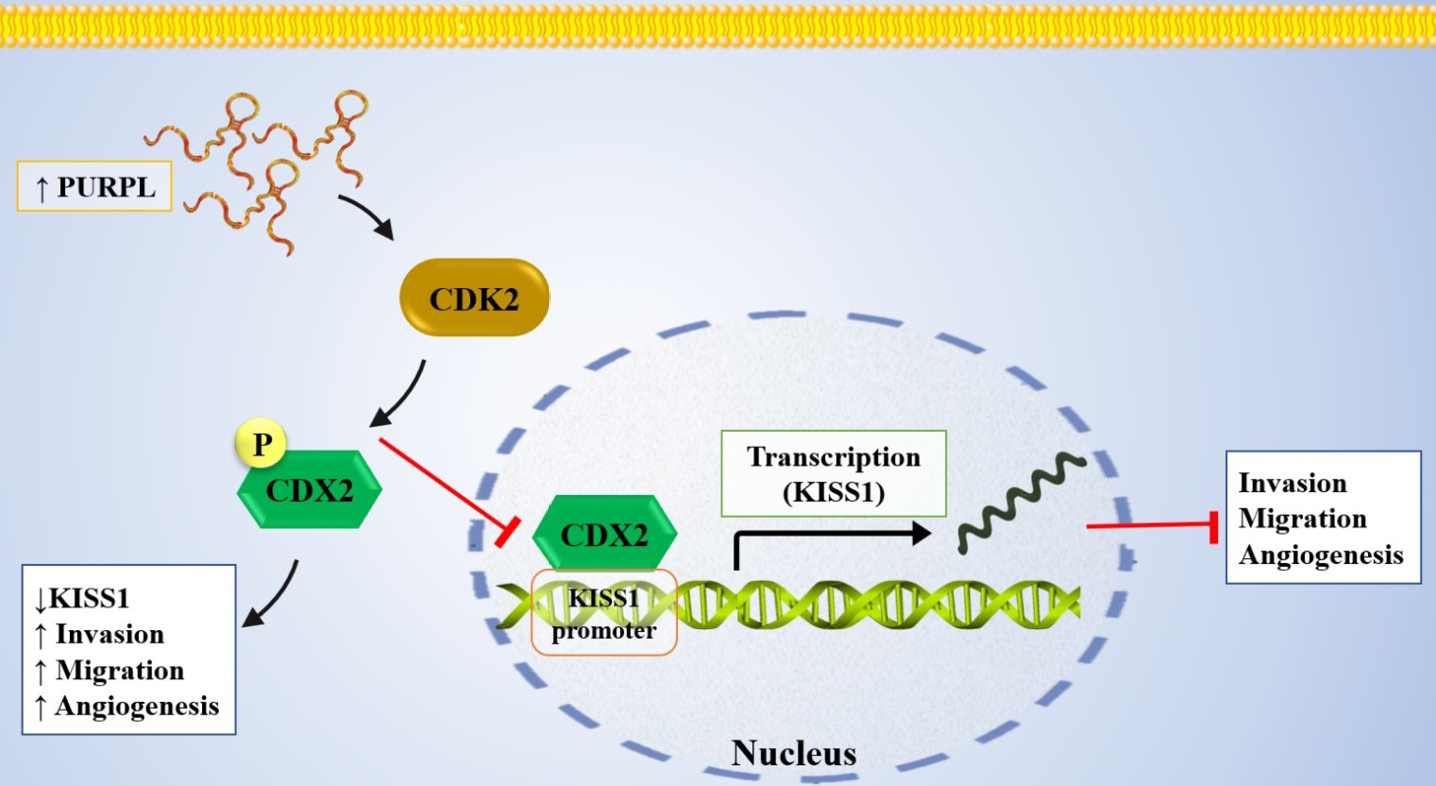
Fig. 6.
Molecular schematic illustrating the role of PURPL in gastric cancer. PURPL is upregulated in gastric cancer and promotes CDK2-mediated phosphorylation of CDX2, ultimately reducing the expression of the tumor suppressor KISS1.
.
Molecular schematic illustrating the role of PURPL in gastric cancer. PURPL is upregulated in gastric cancer and promotes CDK2-mediated phosphorylation of CDX2, ultimately reducing the expression of the tumor suppressor KISS1.
Considering its regulatory impact on key cancer-related pathways, PURPL has emerged as a potential therapeutic target, and the following section focuses on how its knockdown can enhance chemosensitivity in cancer cells.
PURPL knockdown as a strategy to enhance chemosensitivity
Chemotherapy represents one of the most important cancer treatments, aimed at eradicating rapidly dividing tumor cells and controlling disease progression.53 However, patient responses to chemotherapy may run the gamut even within tumors of the same type due to tumor heterogeneity, genetic and epigenetic differences, and the influence of the tumor microenvironment.53 A major challenge in this context is chemoresistance, which enables tumor cells to survive despite drug treatment. Key mechanisms include drug efflux, enhanced DNA repair, evasion of apoptosis, and microenvironment-mediated protection.54 Overcoming these barriers can be facilitated by several new strategies that are being developed to enhance therapeutic efficacy and make cancer treatment more personalized, including targeted drug delivery, nanocarriers, and combination therapies.54 Specifically, lncRNAs have been shown to affect the efficacy of chemotherapeutic agents, since their modulation enhances drug-induced apoptosis and suppresses the growth of tumor cells. In a study directed by Berhane et al, attention has been turned to the role of lncRNA PURPL in HCC and its effect on response to chemotherapy.47 PURPL is a p53-responsive lncRNA that is significantly upregulated in HCC tissues compared with normal liver. Functional experiments using antisense oligonucleotides and siRNA-mediated knockdown in HCC cell lines, such as HepG2 and SK-hep-1, revealed that suppression of PURPL reduces cell proliferation and induces apoptosis, pointing out its critical role in sustaining tumor cell survival.47 Furthermore, knockdown of PURPL also sensitizes HCC cells to doxorubicin. They suggested that PURPL interacts with the p53 signaling pathway modulate apoptotic and cell cycle-related processes in response to chemotherapy. These findings indicate that targeting PURPL could serve as a novel therapeutic strategy to enhance chemosensitivity and overcome drug resistance in liver cancer (Table 1).47
In another study, by Kaller et al, the role of PURPL in CRC and its influence on chemotherapeutic response was investigated. PURPL expression was indeed significantly upregulated in CRC cell lines, and patient-derived samples with functional p53.48 Functional experiments via siRNA-mediated knockdown and CRISPR/Cas9-mediated promoter deletion in CRC cell lines like HCT116 demonstrated that downregulation of LINC01021 promotes apoptosis and inhibits colony formation by modulating tumor cell survival.48 More significantly, it elicits an increased sensitivity of CRC cells to chemotherapeutic agents such as doxorubicin and 5-fluorouracil.48 Mechanistically, while LINC01021 is a p53-inducible target, depleting it failed to alter the expression and activity of p53 itself but seemed to modulate the p53-dependent downstream gene programs, thus influencing the degree of drug-induced apoptosis (Table 1).48
Conclusion and future perspectives
Over the past years, comprehensive studies have unraveled the diverse regulatory functions of lncRNAs in cancer biology. Among these, PURPL has emerged as a significant intergenic lncRNA that is involved in a number of cellular functions, such as miRNA network modulation, autophagy regulation, genomic stability, etc. Its possible oncogenic relevance is supported by the fact that PURPL has been shown to exhibit upregulated expression in a number of cancers, including colorectal, liver, gastric, and melanoma cancers. Although current studies indicate that PURPL plays multiple regulatory roles, these findings are largely derived from preliminary experimental evidence and have yet to be thoroughly validated in human tissues or clinical settings. Moreover, PURPL’s functions are highly context-dependent, influenced by factors such as p53 status and cell type, which can lead to diverse or even opposing roles. The lack of in-depth mechanistic studies on its molecular interactions and epigenetic regulation further limits a comprehensive understanding of PURPL’s actual functions. These considerations underscore the need for future studies employing integrated mechanistic and clinical approaches.
Most notably, knockdown of PURPL alone significantly reversed cancer progression, which is achieved by the suppression of cell proliferation and the induction of apoptosis. Moreover, its suppression enhanced the cytotoxic effects of chemotherapeutic agents, resulting in improved drug responsiveness and overall treatment efficacy. While there is currently no direct evidence to confirm that PURPL itself drives chemoresistance, knockdown indeed seems to improve drug responsiveness, hence providing a therapeutic advantage in combination with conventional chemotherapy.
Research Highlights
What is the current knowledge?
-
PURPL, a key lncRNA, is crucial in gene regulation, p53 pathway control, autophagy, and chromosomal stability.
-
It acts as a miRNA sponge, guide, and scaffold, integrating multiple cellular signaling networks.
What is new here?
Competing Interests
The authors declare no competing interest
Data Availability Statement
No datasets were generated or analyzed during the current study.
Declaration of AI-assisted Tools in the Writing Procedure
ChatGPT (OpenAI) and Grammarly were used solely for language editing, grammar checking, and improving the clarity of the manuscript; however, all scientific content and interpretations are solely the authors’ responsibility.
Ethical Approval
This study is a review article based on previously published research; therefore, no ethical approval or informed consent was required.
Acknowledgements
The authors are thankful for the support of the Immunology Research Center, Tabriz University of Medical Science.
References
- Wang Y, Jiang R, Wang Q, Li Y, Sun Z, Zhao H. Silencing LINC01021 inhibits gastric cancer through upregulation of KISS1 expression by blocking CDK2-dependent phosphorylation of CDX2. Mol Ther Nucleic Acids 2021; 24:832-44. doi: 10.1016/j.omtn.2021.01.025 [Crossref] [ Google Scholar]
- Ferrer J, Dimitrova N. Transcription regulation by long non-coding RNAs: mechanisms and disease relevance. Nat Rev Mol Cell Biol 2024; 25:396-415. doi: 10.1038/s41580-023-00694-9 [Crossref] [ Google Scholar]
- Azadi SS, Safaralizadeh R, Amini M, Hosseinpour Feizi MA, Abdolzadeh S, Najafi S. Investigating the effect of LncRNA DLGAP1-AS2 suppression on chemosensitivity of gastric cancer to chemotherapy. Naunyn Schmiedebergs Arch Pharmacol 2024; 397:7891-903. doi: 10.1007/s00210-024-03130-7 [Crossref] [ Google Scholar]
- Bhat SA, Ahmad SM, Mumtaz PT, Malik AA, Dar MA, Urwat U. Long non-coding RNAs: Mechanism of action and functional utility. Noncoding RNA Res 2016; 1:43-50. doi: 10.1016/j.ncrna.2016.11.002 [Crossref] [ Google Scholar]
- Yan H, Bu P. Non-coding RNA in cancer. Essays Biochem 2021; 65:625-39. doi: 10.1042/EBC20200032 [Crossref] [ Google Scholar]
- Gencel-Augusto J, Wu W, Bivona TG. Long non-coding RNAs as emerging targets in lung cancer. Cancers 2023; 15:3135. doi: 10.3390/cancers15123135 [Crossref] [ Google Scholar]
- Tao H, Yang J-J, Zhou X, Deng Z-Y, Shi K-H, Li J. Emerging role of long noncoding RNAs in lung cancer: Current status and future prospects. Respir Med 2016; 110:12-9. doi: 10.1016/j.rmed.2015.10.006 [Crossref] [ Google Scholar]
- Gao N, Li Y, Li J, Gao Z, Yang Z, Li Y. Long non-coding RNAs: the regulatory mechanisms, research strategies, and future directions in cancers. Front Oncol 2020; 10:598817. doi: 10.3389/fonc.2020.598817 [Crossref] [ Google Scholar]
- Coan M, Haefliger S, Ounzain S, Johnson R. Targeting and engineering long non-coding RNAs for cancer therapy. Nat Rev Genet 2024; 25:578-95. doi: 10.1038/s41576-024-00693-2 [Crossref] [ Google Scholar]
- Chen X, Sun Y, Cai R, Wang G, Shu X, Pang W. Long noncoding RNA: multiple players in gene expression. BMB Rep 2018; 51:280. doi: 10.5483/BMBRep.2018.51.6.025 [Crossref] [ Google Scholar]
- Soltani R, Amini M, Mazaheri Moghaddam M, Jebelli A, Ahmadiyan S, Bidar N. LncRNA DLGAP1-AS2 overexpression associates with gastric tumorigenesis: a promising diagnostic and therapeutic target. Mol Biol Rep 2022; 49:6817-26. doi: 10.1007/s11033-021-07038-w [Crossref] [ Google Scholar]
- Li XL, Subramanian M, Jones MF, Chaudhary R, Singh DK, Zong X. Long noncoding RNA PURPL suppresses basal p53 levels and promotes tumorigenicity in colorectal cancer. Cell Rep 2017; 20:2408-23. doi: 10.1016/j.celrep.2017.08.041 [Crossref] [ Google Scholar]
- Kazemzadeh M, Safaralizadeh R, Mokhtarzadeh AA, Feizi MAH. Understanding the dysregulation of PURPL, a novel long intergenic noncoding RNA, in thyroid cancer progression. Human Gene 2025; 46:201499. doi: 10.1016/j.humgen.2025.201499 [Crossref] [ Google Scholar]
- Wang H, Guo M, Wei H, Chen Y. Targeting p53 pathways: Mechanisms, structures and advances in therapy. Signal Transduct Target Ther 2023; 8:92. doi: 10.1038/s41392-023-01347-1 [Crossref] [ Google Scholar]
- Hill RJ, Bona N, Smink J, Webb HK, Crisp A, Garaycoechea JI. p53 regulates diverse tissue-specific outcomes to endogenous DNA damage in mice. Nat Commun 2024; 15:2518. doi: 10.1038/s41467-024-46844-1 [Crossref] [ Google Scholar]
- Tatavosian R, Donovan MG, Galbraith MD, Duc HN, Szwarc MM, Joshi MU. Cell differentiation modifies the p53 transcriptional program through a combination of gene silencing and constitutive transactivation. Cell Death Differ 2023; 30:952-65. doi: 10.1038/s41418-023-01113-4 [Crossref] [ Google Scholar]
- Ono W, Hayashi Y, Yokoyama W, Kuroda T, Kishimoto H, Ito I. The nucleolar protein Myb-binding protein 1A (MYBBP1A) enhances p53 tetramerization and acetylation in response to nucleolar disruption. J Biol Chem 2014; 289:4928-40. doi: 10.1074/jbc.M113.474049 [Crossref] [ Google Scholar]
- Ajoolabady A, Pratico D, Bahijri S, Tuomilehto J, Uversky VN, Ren J. Hallmarks of cellular senescence: biology, mechanisms, regulations. Exp Mol Med 2025; 57:1482-91. doi: 10.1038/s12276-025-01480-7 [Crossref] [ Google Scholar]
- Rossi C, Venturin M, Gubala J, Frasca A, Corsini A, Battaglia C. PURPL and NEAT1 long non-coding RNAs are modulated in vascular smooth muscle cell replicative senescence. Biomedicines 2023; 11:3228. doi: 10.3390/biomedicines11123228 [Crossref] [ Google Scholar]
- Chen X, Agustinus AS, Li J, Dibona M, Bakhoum SF. Chromosomal instability as a driver of cancer progression. Nat Rev Genet 2025; 26:31-46. doi: 10.1038/s41576-024-00761-7 [Crossref] [ Google Scholar]
- Bakhoum SF, Cantley LC. The multifaceted role of chromosomal instability in cancer and its microenvironment. Cell 2018; 174:1347-60. doi: 10.1016/j.cell.2018.08.027 [Crossref] [ Google Scholar]
- Malakar P. Long noncoding RNA, PURPL is associated with aneuploidy and its magnitude of expression level is dependent on P53 status. Front Cell Dev Biol 2024; 12:1410308. doi: 10.3389/fcell.2024.1410308 [Crossref] [ Google Scholar]
- Alshahrani SH, Al-Hadeithi ZS, Almalki SG, Malviya J, Hjazi A, Mustafa YF. LncRNA-miRNA interaction is involved in colorectal cancer pathogenesis by modulating diverse signaling pathways. Pathol Res Pract 2023; 251:154898. doi: 10.1016/j.prp.2023.154898 [Crossref] [ Google Scholar]
- Hejazi M, Heshmat R, Shafiee G, Larijani B, Mokhtarzadeh AA, Ebrahimi V. The Interplay Between lncRNAs–microRNAs Network Dysregulation and Cellular Hallmarks of Thyroid Cancer. Cancers 2025; 17:3373. doi: 10.3390/cancers17203373 [Crossref] [ Google Scholar]
- Ahmadpour Youshanlui M, Yari A, Bahojb Mahdavi SZ, Amini M, Baradaran B, Ahangar R. BRD4 expression and its regulatory interaction with miR-26a-3p, DLG5-AS1, and JMJD1C-AS1 lncRNAs in gastric cancer progression. Discov Oncol 2024; 15:356. doi: 10.1007/s12672-024-01230-7 [Crossref] [ Google Scholar]
- Chow JT-S, Desjardins A, Lee DK, Grigore IA, Lee L, Fu NJ. A microRNA CRISPR screen reveals microRNA-483-3p as an apoptotic regulator in prostate cancer cells. Cell Death Dis 2025; 16:752. doi: 10.1038/s41419-025-08098-7 [Crossref] [ Google Scholar]
- Hejazi M, Bahojb Mahdavi SZ, Abedimanesh S, Heshmat R, Larijani B, Mokhtarzadeh AA. LINC00162 silencing enhances sorafenib sensitivity and inhibits thyroid cancer cells progression through modulation of MAPK signaling and apoptosis. Sci Rep 2025; 15:29726. doi: 10.1038/s41598-025-12805-x [Crossref] [ Google Scholar]
- Hejazi M, Jafari T, Yari A, Heshmat R, Larijani B, Ahvaz S. A novel long non-coding RNA, PICSAR, promotes thyroid cancer progression through the hsa-miR-320A/hsa-miR-485/RAPGEFL1 axis. Med Oncol 2025; 42:448. doi: 10.1007/s12032-025-02987-9 [Crossref] [ Google Scholar]
- Wang Y, Chen T, Huang H, Jiang Y, Yang L, Lin Z. miR-363-3p inhibits tumor growth by targeting PCNA in lung adenocarcinoma. Oncotarget 2017; 8:20133. doi: 10.18632/oncotarget.15448 [Crossref] [ Google Scholar]
- Zhang R, Guo X, Zhao L, He T, Feng W, Ren S. Abnormal expressions of PURPL, miR-363-3p and ADAM10 predicted poor prognosis for patients with ovarian serous cystadenocarcinoma. J Cancer 2023; 14:2908. doi: 10.7150/jca.87405 [Crossref] [ Google Scholar]
- Alabi RO, Farber G, Blobel CP. Intriguing roles for endothelial ADAM10/Notch signaling in the development of organ-specific vascular beds. Physiol Rev 2018; 98:2025-61. doi: 10.1152/physrev.00029.2017 [Crossref] [ Google Scholar]
- Shi Q, Xue C, Zeng Y, Yuan X, Chu Q, Jiang S. Notch signaling pathway in cancer: from mechanistic insights to targeted therapies. Signal Transduct Target Ther 2024; 9:128. doi: 10.1038/s41392-024-01828-x [Crossref] [ Google Scholar]
- Pan Y, Yu Y, Wang X, Zhang T. Tumor-associated macrophages in tumor immunity. Front Immunol 2020; 11:583084. doi: 10.3389/fimmu.2020.583084 [Crossref] [ Google Scholar]
- Kzhyshkowska J, Shen J, Larionova I. Targeting of TAMs: can we be more clever than cancer cells?. Cell Mol Immunol 2024; 21:1376-409. doi: 10.1038/s41423-024-01232-z [Crossref] [ Google Scholar]
- Wang S, Wang J, Chen Z, Luo J, Guo W, Sun L. Targeting M2-like tumor-associated macrophages is a potential therapeutic approach to overcome antitumor drug resistance. NPJ Precis Oncol 2024; 8:31. doi: 10.1038/s41698-024-00522-z [Crossref] [ Google Scholar]
- Wang Y, Jia J, Wang F, Fang Y, Yang Y, Zhou Q. Pre-metastatic niche: formation, characteristics and therapeutic implication. Signal Transduct Target Ther 2024; 9:236. doi: 10.1038/s41392-024-01937-7 [Crossref] [ Google Scholar]
- He F, Ding G, Jiang W, Fan X, Zhu L. Effect of tumor-associated macrophages on lncRNA PURPL/miR-363/PDZD2 axis in osteosarcoma cells. Cell Death Discov 2021; 7:307. doi: 10.1038/s41420-021-00700-z [Crossref] [ Google Scholar]
-
Yi Q, Cui H, Liao Y, Xiong J, Ye X, Sun W. A minor review of microRNA-338 exploring the insights of its function in tumorigenesis. Biomed Pharmacother 2021. 139: 111720. doi: 10.1016/j.biopha.2021.111720.
- Zhang R, He T, Shi H, Yuan C, Wei F, Liu Z. Disregulations of PURPL and MiR-338-3p could serve as prognosis biomarkers for epithelial ovarian cancer. J Cancer 2021; 12:5674. doi: 10.7150/jca.61327 [Crossref] [ Google Scholar]
- Cheng Z, Hong J, Tang N, Liu F, Gu S, Feng Z. Long non-coding RNA p53 upregulated regulator of p53 levels (PURPL) promotes the development of gastric cancer. Bioengineered 2022; 13:1359-76. doi: 10.1080/21655979.2021.2017588 [Crossref] [ Google Scholar]
- Liu S, Ruan Y, Chen X, He B, Chen Q. miR-137: a potential therapeutic target for lung cancer. Front Cell Dev Biol 2024; 12:1427724. doi: 10.3389/fcell.2024.1427724 [Crossref] [ Google Scholar]
- Wu H, Ding X, Hu X, Zhao Q, Chen Q, Sun T. LINC01021 maintains tumorigenicity by enhancing N6-methyladenosine reader IMP2 dependent stabilization of MSX1 and JARID2: implication in colorectal cancer. Oncogene 2022; 41:1959-73. doi: 10.1038/s41388-022-02189-x [Crossref] [ Google Scholar]
-
Yang Y, Jia X, Peng Y, Lu S, Wang C, Zhang H, et al. Phosphorylation of MSX1 controls tumorigenesis and bone development through targeting FBXW7 degradation. Cancer Lett 2025; 217948. doi: 10.1016/j.canlet.2025.217948.
- Liu C, Huang M, Han C, Li H, Wang J, Huang Y. A narrative review of the roles of muscle segment homeobox transcription factor family in cancer. Ann Transl Med 2021; 9:810. doi: 10.21037/atm-21-220 [Crossref] [ Google Scholar]
- Cheng Y, Song Z, Cheng J, Tang Z. JARID2, a novel regulatory factor, promotes cell proliferation, migration, and invasion in oral squamous cell carcinoma. BMC Cancer 2024; 24:793. doi: 10.1186/s12885-024-12457-6 [Crossref] [ Google Scholar]
- Han S, Li X, Wang K, Zhu D, Meng B, Liu J. PURPL represses autophagic cell death to promote cutaneous melanoma by modulating ULK1 phosphorylation. Cell Death Dis 2021; 12:1070. doi: 10.1038/s41419-021-04362-8 [Crossref] [ Google Scholar]
- Berhane T, Holm A, Karstensen KT, Petri A, Ilieva MS, Krarup H. Knockdown of the long noncoding RNA PURPL induces apoptosis and sensitizes liver cancer cells to doxorubicin. Sci Rep 2022; 12:19502. doi: 10.1038/s41598-022-23802-9 [Crossref] [ Google Scholar]
- Kaller M, Götz U, Hermeking H. Loss of p53-inducible long non-coding RNA LINC01021 increases chemosensitivity. Oncotarget 2017; 8:102783. doi: 10.18632/oncotarget.22245 [Crossref] [ Google Scholar]
- Jalali P, Shahmoradi A, Samii A, Mazloomnejad R, Hatamnejad MR, Saeed A. The role of autophagy in cancer: from molecular mechanism to therapeutic window. Front Immunol 2025; 16:1528230. doi: 10.3389/fimmu.2025.1528230 [Crossref] [ Google Scholar]
- Pandey A, Goswami A, Jithin B, Shukla S. Autophagy: The convergence point of aging and cancer. Biochem Biophys Rep 2025; 42:101986. doi: 10.1016/j.bbrep.2025.101986 [Crossref] [ Google Scholar]
- Fares J, Fares MY, Khachfe HH, Salhab HA, Fares Y. Molecular principles of metastasis: a hallmark of cancer revisited. Signal Transduct Target Ther 2020; 5:28. doi: 10.1038/s41392-020-0134-x [Crossref] [ Google Scholar]
- Dai X, Xi M, Li J. Cancer metastasis: molecular mechanisms and therapeutic interventions. Mol Biomed 2025; 6:20. doi: 10.1186/s43556-025-00261-y [Crossref] [ Google Scholar]
- Ulukaya E, Karakas D, Dimas K. Tumor chemosensitivity assays are helpful for personalized cytotoxic treatments in cancer patients. Medicina 2021; 57:636. doi: 10.3390/medicina57060636 [Crossref] [ Google Scholar]
- Davodabadi F, Sajjadi SF, Sarhadi M, Mirghasemi S, Hezaveh MN, Khosravi S. Cancer chemotherapy resistance: Mechanisms and recent breakthrough in targeted drug delivery. Eur J Pharmacol 2023; 958:176013. doi: 10.1016/j.ejphar.2023.176013 [Crossref] [ Google Scholar]